PLS
622 Plant Physiology I Wednesday, October 04, 2006
Reproductive
development:
Objectives:
1) Investigate the determinacy of the
conversion from the vegetative, to the inflorescent, to the floral meristem.
2) Examine what we know regarding the
molecular control over flowering time in arabidopsis.
Transition
to flowering II:
So far we have
examined how environmental cues are perceived by the plant and how that
perception leads to the production of elicitors that alter the fate of the
shoot apical meristem, changing it from a vegetative meristem to an
inflorescence meristem and finally to a floral meristem. We will next look at
how the change from a vegetative to a reproductive meristem is orchestrated.
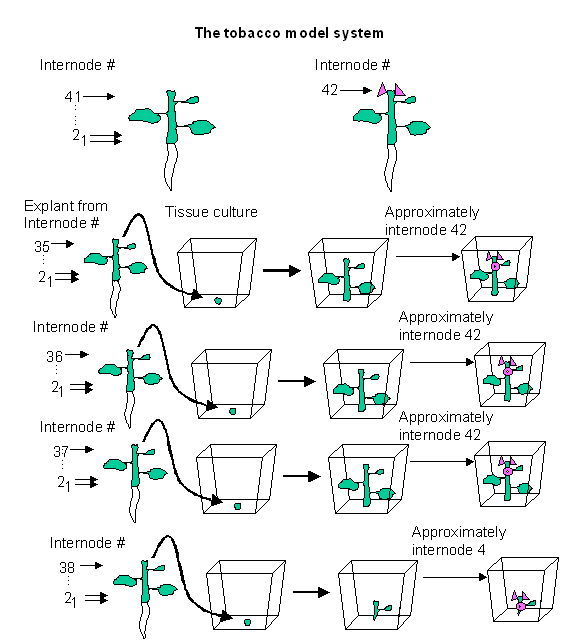
It
is useful to study plant anatomy comparatively. This serves to simplify the
seemingly endless variations of morphology down to basic anatomical features
from which many plant organs can be said to arise. There are five fundamental
aspects of plant growth that can be employed to study plant anatomy comparatively.
Variability in internode length; phyllotaxy (placement and arrangement
of parts on an organ); and determinacy
dictate the shape a plant organ can assume while organ identity determines the function the organ(s) under study
will perform. Lastly, the programmed
destruction of organ parts through programmed cell death (PCD, a.k.a.
apoptosis) determines the shape and function a plant part finally attains. All
of these features play a role in the transition from a vegetative meristem to
an inflorescence meristem. The switch from vegetative to reproductive life
cycles is neither subtle nor simple but involves changes in gene expression
that affects the nature and identity of the structures produced from the stem
cells of the meristem. So, although the stem cells of the vegetative meristem
are as undifferentiated as the stem cells of the inflorescence or floral
meristems, the vegetative meristem is capable of producing only stem, leaves,
and vegetative buds while the inflorescence meristem produces floral bracts,
inflorescence branches, and flowers. Finally, the switch from a vegetative
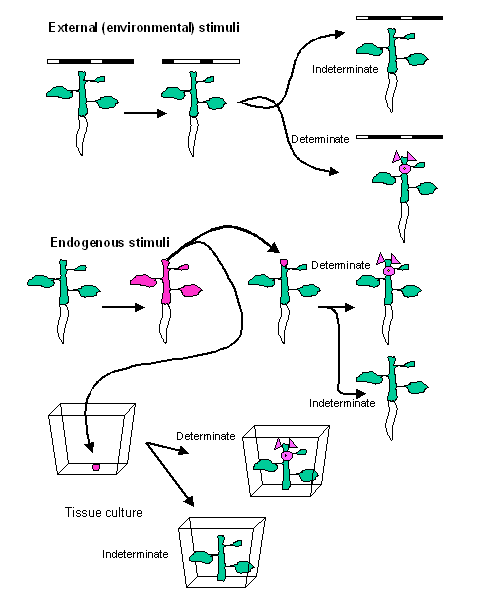
meristem to an inflorescence meristem is often determinant but not always! This
means that, in most plants, even if the stimuli triggering the transition from
vegetative to reproductive growth are removed, the inflorescence meristem is
irrevocably committed to producing flowers and cannot revert to the vegetative
mode. This is relatively easy to test in plants that have an external
environmental cue they use to switch from vegetative to reproductive
growth…simply remove the inductive stimulus. It is more difficult when the
stimulus is endogenous but it can still be done using elaborate grafting or
tissue culture techniques.
However, there
are some species that can revert to the vegetative mode following induction of
the reproductive life cycle if they are moved to non-inductive environments. In
some species of Petunia, if the plants are removed from inductive short days to
non-inductive long days the phenomenon of floral
reversion converts the meristem back to the vegetative mode. This can be
quite dramatic since occasionally, even plants that have gone so far as to
initiate carpels and ovules will revert and the vegetative meristem will
commence elongation from the carpel! Floral reversion is one manner of having a
floral meristem turn back to vegetative growth. Partial flowering is a second manner of having floral and
vegetative meristem activity interspersed along a plant axis. In this case some
meristems that would otherwise have developed into flowers are kept in the
vegetative mode while neighboring meristems produce flowers. Plants given
sub-optimal stimulus for transition to flowering often exhibit partial
flowering.
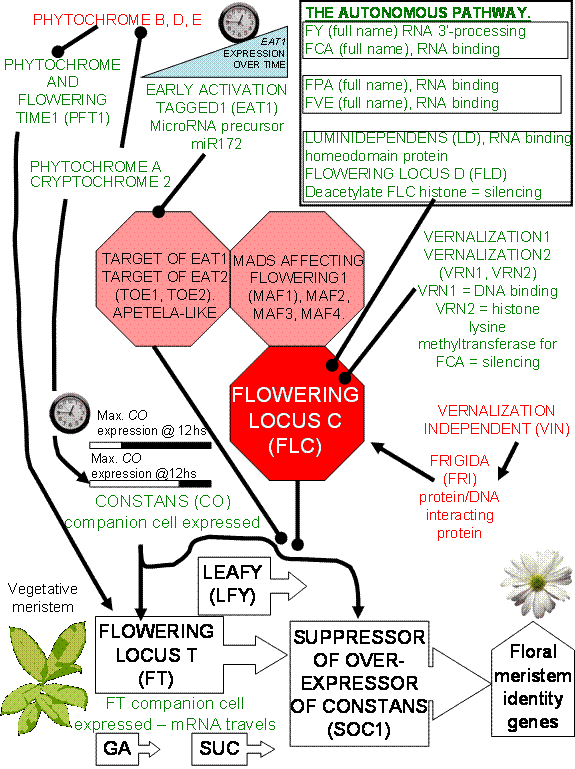
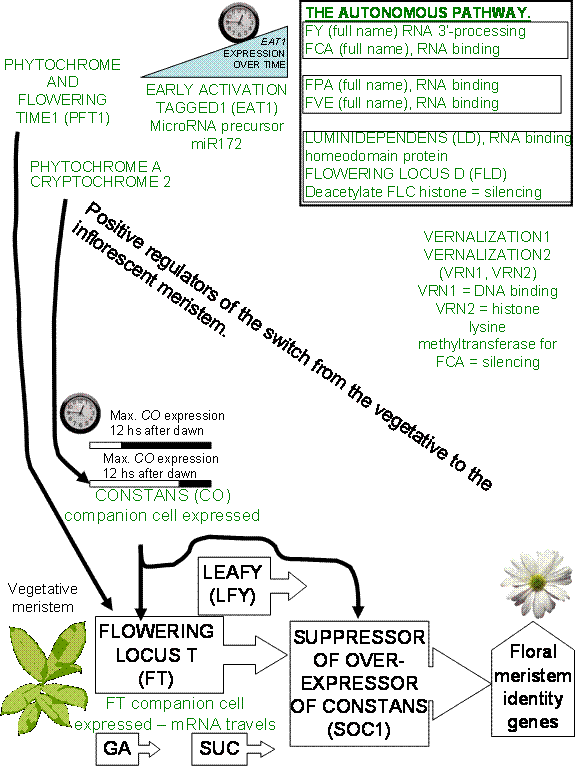
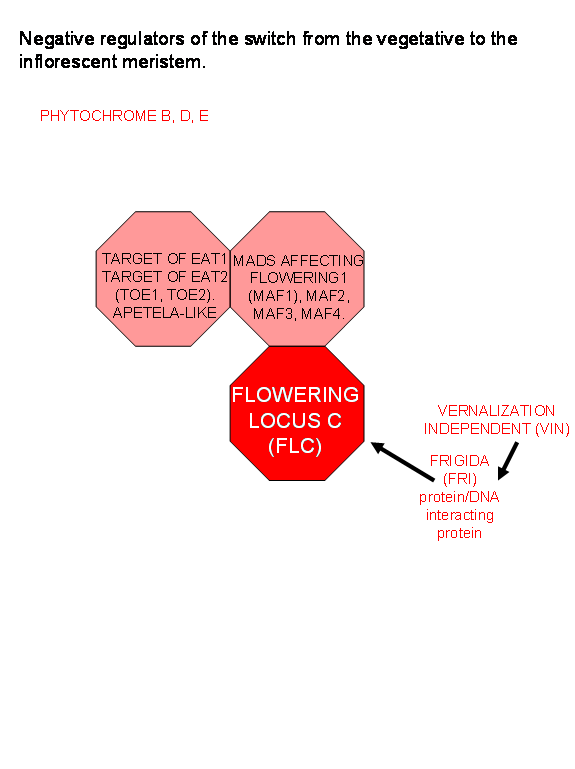
The
control of whether a meristem remains vegetative or switches to an
inflorescence meristem appears to be controlled by several different pathways,
involving many different genes. Mutant analysis of the ever increasing number
of mutants causing late flowering were shown to sort into 3 different epistatic
groups. This means that there are probably at least 3 independent paths
exerting control over the time to flowering.
If you recall from the previous
lecture, one of the first elicitors signaling the transition to flowering was
the mobilization of starch into glucose, which was subsequently used to
synthesize sucrose which was transported to the shoot apical meristem and the
root. Mutants affected in their ability to either make starch (phosphoglucomutanse deficient (pgm)) or their ability to hydrolyze it (starch overproducer (sop)) should therefore, encounter
difficulties in entering the reproductive mode. If grown under continuous light
the mutants flower at the same time as wild type. However, as day length is
decreased they flower increasingly later than the wild type. Vernalization
completely restores the wild type flowering regime to these mutants.
GA
deficient mutants commence flowering at the same time as wild type under long
days but are absolutely incapable of flowering under short days without exogenous
GA. This demonstrates that GA is necessary for the transition to flowering in
arabidopsis. Abscisic acid deficient mutants flower earlier than wild type
demonstrating that
Two
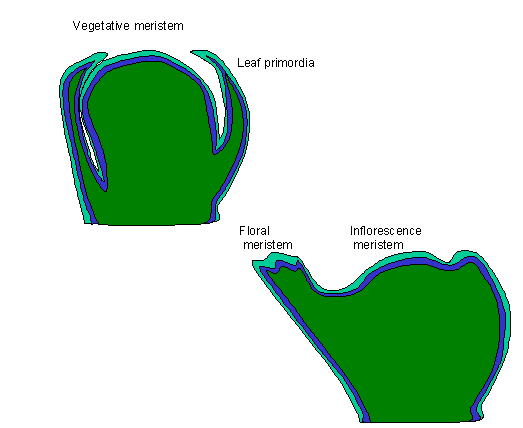
types of genes are said to influence inflorescence development; 1) meristem identity genes and, 2) organ identity genes. The former
generally affects the first four aspects of plant growth (internode length; phyllotaxy;
determinacy; and organ identity) while the latter affects the fate of organ
primordia and therefore directly
determines the identity and existence of the organ. All normal plants commence
post-establishment life with a vegetative stage of development where leaf
primordia and lateral branches are produced on the periphery of an apical
meristem. Upon induction of flowering, this vegetative meristem undergoes a transition to an inflorescence meristem typified by the
production of, 1) shorter internodes;
2) hairy stems; 3) small bracts; and 4) decreased plastochron durations. Finally, the inflorescence meristem produces
flower meristems peripherally and
continuously (indeterminate growth) or at the apical meristem once
(determinate) which prevents further growth on that axis. Hence, inflorescence
development can be thought of as sequential with the vegetative meristem
producing the inflorescence meristem which leads to the floral meristem. The
switch from the vegetative to the inflorescent meristem is under considerable
environmental and developmental control.
To make life
even more complex, the determination of floral primordia and the determination
of floral organ primordia are also separate events. There are many mutants (apetala2, agamous, pistillata, and superman) that affect the order and
identity of the floral organ produced without affecting the determination of
the flower itself. That is to say, these mutants initiate the production of a
flower, exhibiting such floral traits as shortened internodes and whorled
arrangement of organs, but the floral organs themselves are independently
determined and in the mutants have their identity altered.
A
mutant in a gene important in determining floral meristem identity has been
isolated from arabidopsis. The leafy (lfy) mutation produces shoots in the axils of cauline leaves of the
inflorescence where the first, bractless flowers normally develop. The shoots
become increasingly flower-like as one moves acropetally towards the apex with
male and female organs eventually forming in a spiral phyllotaxis. It appears
as though the lfy gene affects
partitioning of a group of cells (anlagen)
that together will form the bractless floral meristem in wild-type and the
meristem and the cauline leaf primordia in the mutant. The wild-type condition
where the flower meristem develops without a sub-tending bract is thought to be
a derived condition from an ancestor whose flowers did develop bracts. Lfy in the wild-type is thought to
recruit all the cells of the anlagen into the floral meristem leaving none for
the bract primordium, possibly leading to a faster rate of floral development.
Additionally, the terminal flower (tfl)
mutant of arabidopsis produces a flower at the apex of the apical meristem
thereby preventing any further growth on that axis. The wild-type plants have
indeterminate growth whereby the floral meristem is always located peripherally
on the inflorescent meristem permitting continued (indeterminate) growth and
flower production. The tfl mutant
affects the determinacy of the inflorescence meristem changing it to
determinate. The gene product of wild-type terminal flower must somehow be
antagonistic to floral development since whenever it is normal and active, it
represses the production of a floral meristem in the apex.
There are many
different genes that have been found to affect the timing of the transition to
flowering. These mutations can be grouped into those delaying flowering and
those enhancing the entry into the reproductive stage of the life cycle. Null
mutants of the CONSTANS (CO) and GIGANTEA (GI) genes (co and gi, respectively) hardly respond to vernalization or long days. At
the other end of the spectrum the mutants terminal
flower (tfl), and early flowering1 (elf1) genes, cause early entry into the flowering phase of the life
cycle in response to the same environmental cues as the wild type. Probably the
most severe mutants affecting entry into the reproductive stage of the plant
life cycle are those in the EMBRYONIC
FLOWERING (EMF) gene. The emf mutant proceeds directly to
inflorescence growth following the completion of germination.