PLS 622: Plant
Physiology I, Friday, October 20, 2006 Reproductive
development:
Self-incompatibility:
Objectives:
An understanding of the current state of knowledge of sporophytic and
gametophytic self-incompatibility.
In the last lecture we investigated
pollen development and pollen tube growth through transmitting styles. In pollination,
the pollen grains traveled via animals or air currents to other flowers in the
vicinity or simply fell onto the stigma of the same flower from which the
pollen was produced (self-fertilization). Regardless, the grains must alight on
a stigma from a flower of the same species in order to have a chance of passing
on the genetic information they contain to the next generation. Some flowers
are capable of fertilizing their own ova with the pollen produced on adjacent
anthers. Some species even enhance this self-fertilization by pollinating the
stigma prior to the flower opening (cleistogamy)
(e.g. arabidopsis). Other species are not capable of self-fertilization and are
termed “self-incompatible”. It is this phenomenon and the mechanisms behind it
that we are going to talk about in this lecture.
Self-incompatibility:
There are two major self-incompatibility (SI) systems
distinguished by sporophytic or gametophytic determination of the
self-incompatible reaction. Both are determined by the S-locus, a single,
multiallelic locus that imparts to pollen a specific phenotype recognized by
the stigma and style of potential
receptor plants. In the gametophytic system, the phenotype of the pollen is
determined by the gamete’s genotype. In the sporophytic system, the phenotype
of the pollen is determined by the diploid parent’s genotype. This difference
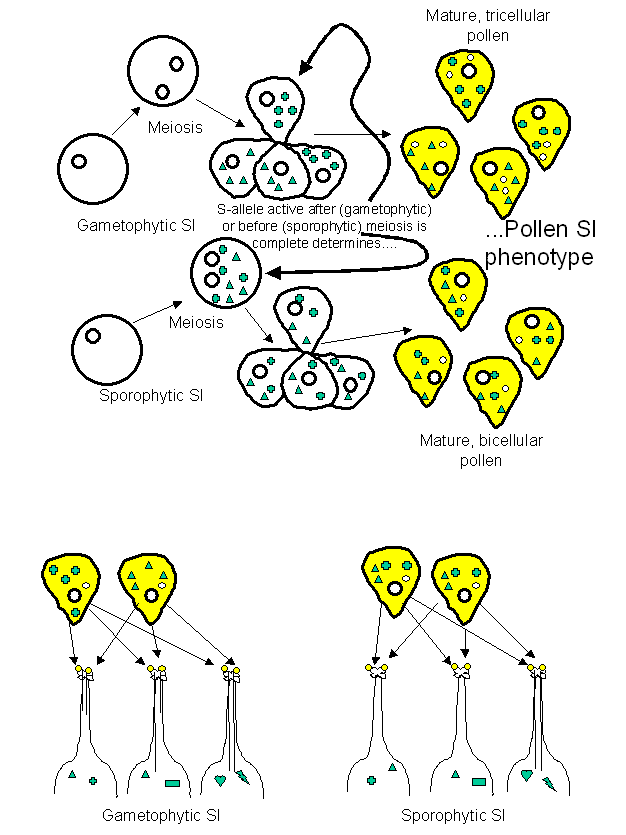
between the two systems has been elegantly explained by assuming that gamete
phenotype is determined by factors expressed after (gametophytic system) or before (sporophytic system) the advent of meiosis. If the S-allele becomes
active prior to meiosis then the gametes obtain and retain both
self-incompatibility factors present in the paternal genotype (sporophytic). If
the S-allele is silent until after meiosis is complete, then only one of the
paternal incompatibility factors is present per gamete (gametophytic) (Fig. 1).
The gametophytic
self-incompatibility system:
In this system fertilization does not occur any time
the S-allele of the gamete matches an active S-allele of the plant to be
fertilized. Typically, the pollen complete germination on the stigma but, at
some point as the pollen tube traverses the transmitting tissue, its progress
is terminated and the pollen tube dies, probably through uptake of RNases
present on the stigma and in the style (Fig. 1).
Figure
1: The proposed mechanism leading to the development of the gametophytic- or
sporophytic-self-incompatibility systems. The green symbols are the proteins
produced from the “S-alleles” (really S-haplotype genes; see below).
The sporophytic
self-incompatibility system:
This system is dependent on the ability of the stigma and
style to differentiate between pollen produced by the same plant or genetically
related plant and pollen from a plant, of the same species, that is not
genetically related. Typically this inhibition is rapid resulting in the
inability of the genetically related pollen to complete germination on the
stigma and/or the inability of the protruded pollen tube to penetrate the
papillar cell wall. The phenotype of the pollen is determined sporophytically
by the diploid genome of the parent plant (Fig. 1). Typically, in the Brassicaceae, the stigma becomes
self-incompatible only 1 day prior to the flower opening.
Mechanisms of
self-incompatibility:
There is a wide variety of alleles associated with
self-incompatibility (upwards of 60 identified in Brassica oleracea) and there are codominant and/or dominant allelic
interactions that determine the intensity of the self-incompatibility response
manifested. This information, combined with the realization that there are at
least three genes responsible for the SI response, has led to the adoption of
the term “S haplotype” rather than
the classical “S-allele” to refer to the inherited information determining this
complex response. Nevertheless, the term “S-allele” is quite entrenched in the
literature and we will continue to use these two terms interchangeably. There
are two major recognized intensities of S-haplotype responses distinguished
based on the number of successful self-pollen germinations occurring in
heterozygous plants. Class I responses
permit the germination and successful pollen tube development of between 0 and
10 per self-pollinated stigma while Class
II responses permit between 10 and 30.
In contrast to previous belief, there are three tightly
linked, transcribed genes that participate in determining self-incompatibility
and hence, the S-locus in both Sporophytic and Gametophytic
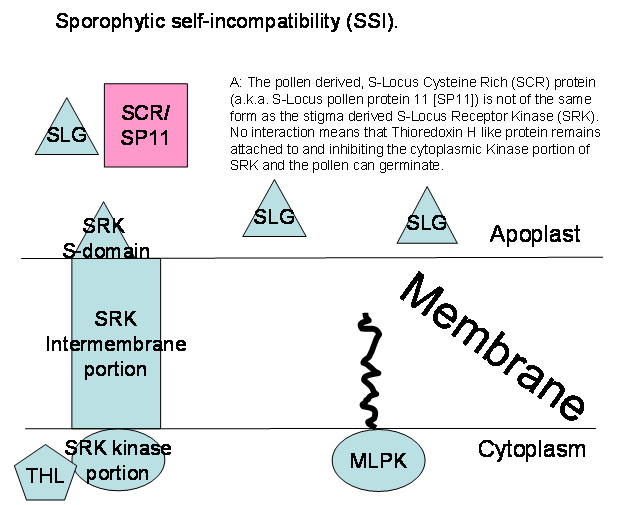
self-incompatibility. One gene is the S-locus glycoprotein (SLG) which is a stigma-transcribed gene
producing an extracellularly excreted protein that acts as an assistant
receptor protein in the sporophytic self-incompatibility response. The second
is highly related in amino-acid homology to SLG over that part of its length
associated with the receptor portion. In addition to the receptor, it also
encodes a membrane spanning domain, and a cytosolic protein kinase domain with
phosphorylating activity (a serine/threonine kinase). This is the S-locus receptor kinase (SRK for S-locus
Receptor Kinase). This membrane associated receptor is only present in the
stigma. The third is the S-locus pollen protein
11 (SP11) or the S-locus Cysteine-Rich protein; SCR), two names for the
same protein. The SP11 gene exhibits
pollen specific gene expression. This protein is deposited in the pollen coat
and leaches out onto the stigma upon the pollen binding to it. The SRK
determines the S-haplotype specificity of the stigma while the SCR/SP11
determines the S-haplotype specificity of the pollen. SLG along with SRK form a
high-affinity receptor in the membrane of the stigma for SCR/SP11. SLG is not
necessary for SRK to bind SCR/SP11 (resulting in a Class II response) but in
those species where it is functional, it can heighten the stringency of the SI
response to a Class I response.
It is currently thought that, at least in the
Brassicaceae, the SLG arose by
duplication of the SLK and selective
deletion of exons and introns to leave a highly expressed (very active
promoter) gene producing a secreted protein, SLG. The SLK has an extracellular
domain that is thought to interact with the SLG to enhance pollen
incompatibility.
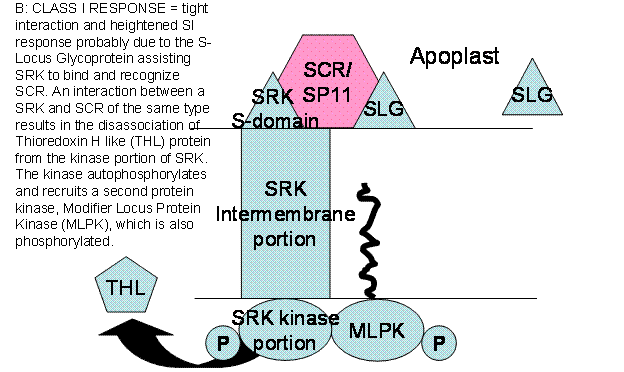
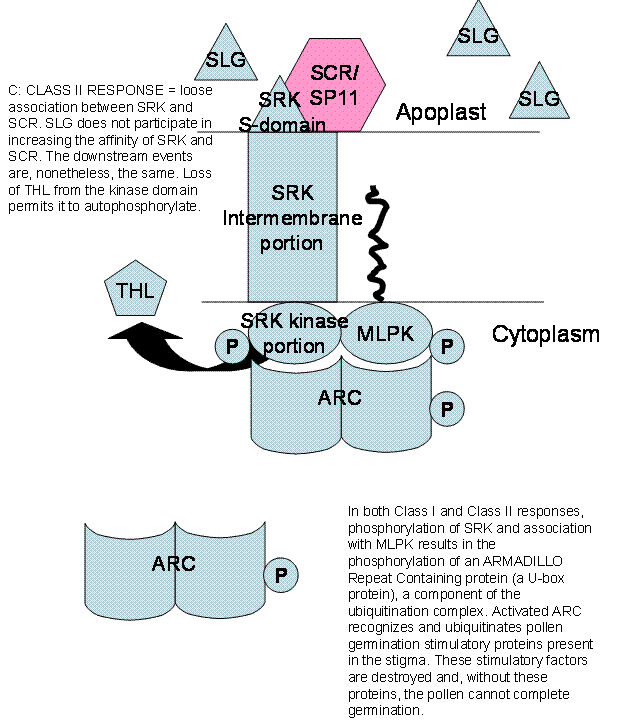
The SRK is usually inhibited from
autophosphorylating itself on the cytoplasmic face of the membrane by a
Thioredoxin H-like protein (THL) bound to it. If the SRK is not phosphorylated
it is inactive. In self-incompatible plants, SRK (with or without the help of
SLG) binds SRC/SP11 from the incompatible pollen and THL immediately
disassociates from the kinase domain. Autophosphorylation activates SRK which
recruits another kinase, Modifier Locus Protein Kinase (MLPK) which together
activate an Armadillo Repeat Containing Protein (ARC). ARC recognizes pollen
growth promoting proteins, poly-ubiquitinates them and has them destroyed via
the 26S proteasome.
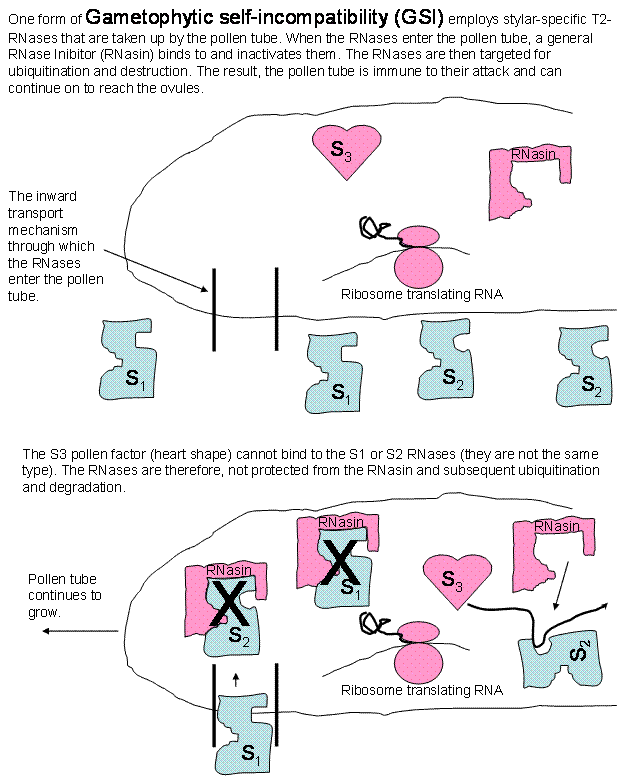
In Gametophytic SI, the Style produces RNases
that are capable of entering the pollen tube. In compatible interactions, once
the RNases are in the tube they are immediately inhibited by a pollen-produced
RNase inhibitor (RNasin) and then ubiquitinated and destroyed. In incompatible
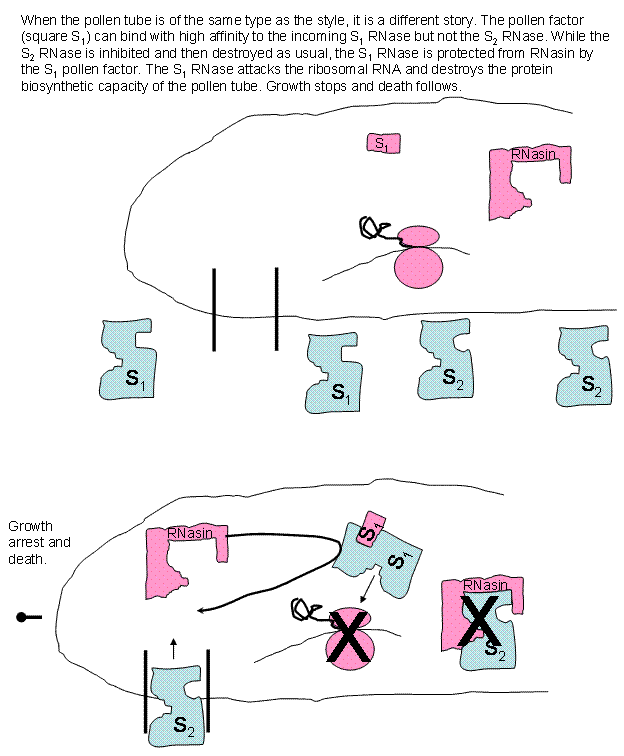
interactions, at least one of the RNases, upon entering the tube, is bound by a
pollen factor that protects the RNase from the RNasin. The protected RNase
attacks the ribosomal RNA and destroys it, the ribosomes, and thus, the protein
production capacity of the tube. Growth is arrested and death soon follows.