
Lecture Twenty one
Lipids -- Fatty Acid Oxidation
Purpose and Outline The purpose of today's lecture/discussion is to give an overview and some detail on specific subjects of fatty acid oxidation unique to plants and of significance in plant metabolism.
__________________________________________________________________________________________________________________
Reading Assignment for the Final Lipid Lecture Discussion:
a) REQUIRED:
1 - Chapter 10, sections 10.10.4 -10.10.5, 10.11 and Chapt. 17 section 17.8 of the Biochemistry & Molecular Biology of Plants class text.
b) OPTIONAL:
1 - Browse, J. 2009. Jasmonate Passes Muster: A Receptor and Targets for the Defense Hormone. Annual Review of Plant Biology 60:183-205.
2 - Koch, et al. 1999. Differential induction of plant volatile biosynthesis in the lima bean by early and late intermediates of the octadecanoid-signaling pathway. Plant Physiol. 121:153-162.
3 - Hamberg, M., A. Sanz, M. J. Rodriguez, A. P. Calvo and C. Castresana. 2003. Activation of the Fatty Acid {alpha}-Dioxygenase Pathway during Bacterial Infection of Tobacco Leaves: FORMATION OF OXYLIPINS PROTECTING AGAINST CELL DEATH. J. Biol. Chem. 278: 51796-51805.
4 - Lorenzo, O. R. Piqueras, J.J. Sanchez-Serrano and R. Solano. 2003. ETHYLENE RESPONSE FACTOR1 Integrates Signals from Ethylene and Jasmonate Pathways in Plant Defense. Plant Cell 15: 165-178.
5 - Pracharoenwattana, I., J.E. Cornah, and S.M. Smith. 2005. Arabidopsis Peroxisomal Citrate Synthase Is Required for Fatty Acid Respiration and Seed Germination. Plant Cell 17: 2037-2048.
__________________________________________________________________________________________________________________
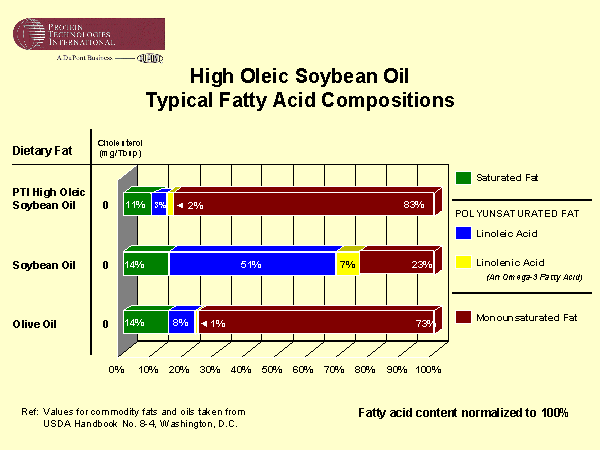
As mentioned in Lecture 20, unsaturated fatty acids are produced by desaturase-catalyzed oxidation of the more reduced saturated fatty acid molecules. Knowledge of fatty acid biosynthesis in plants has led to one of the more significant developments in the biotechnology industry. Vegetable oils (triglyceride) high in polyunsaturated fatty acids are too oxidatively unstable for cooking or spray oil applications. They therefore need to be hydrogenated. Triglycerides high in saturated or monounsaturated fatty acids are oxidatively stable, but monounsaturated fatty acids are much healthier. The best all around cooking and spray oil is olive oil. Recently genetic engineering of soybeans for a reduction of a seed-specific ∆12 desaturase has resulted in the development of a soybean oil with the most favorable fatty acid composition on the market for cooking and spray applications as well as a number of industrial applications. How do you think this was achieved?
Arabidopsis oil synthesis... (Ruuska et al., 2002; Plant Cell 14:1191-1206).
Note Assignment 2 listed on Bb.
We have also already discussed fatty acid oxidation some in the synthesis of many of the monomers of wax, cutin and suberin. The recover of energy in the catabolism of fatty acids from triglycerides is mainly from β-oxidation. This is similar in all organisms and as such is covered in general biochemistry and there is no need to cover the β-oxidation pathway again in lecture here. I would like to emphasize a few points about ß-oxidation in plants such as the differences between plants and animals.
Before b -β-oxidation can begin, fatty acids must 1st be released from triglycerides by a lipase and "activated" to an acyl-CoA e.g.:
18:0 + CoA + ATP ![]() 18:0-CoA+ AMP + PPi = consumption of 2 high energy bounds or 2 ATPs
18:0-CoA+ AMP + PPi = consumption of 2 high energy bounds or 2 ATPs
β-oxidation in plants occurs in glyoxysomes or peroxisomes, but in animals it usually occurs in mitochondria. This results in the lack of reduction of FAD to FADH in the 1st oxidation of the b-β-oxidation cycle in plants.
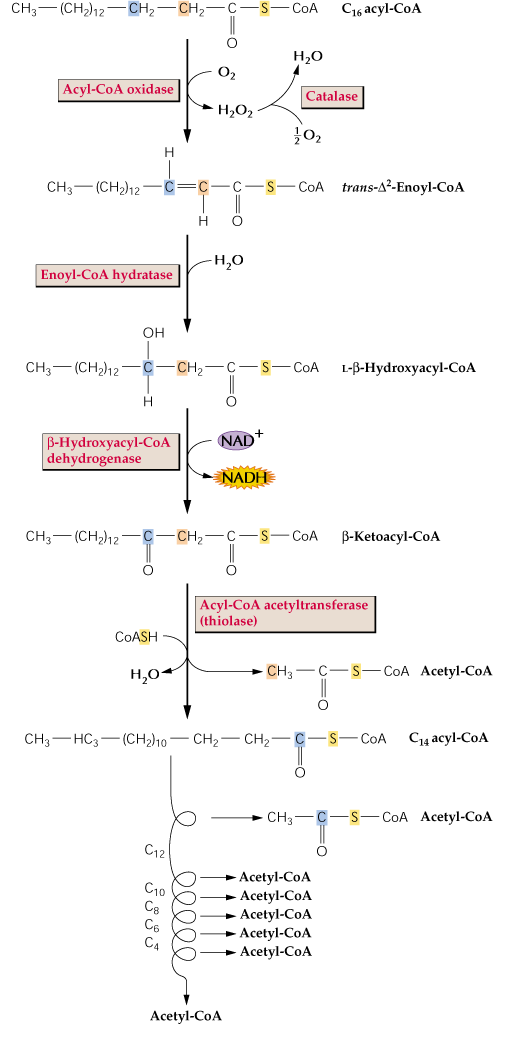
The β-oxidation of polyunsaturated fatty acids (PUFAs) such as 18:3-CoA has several differences compared with β-oxidation of 18:0. We do not have time to cover this except to point out that plants utilize PUFAs somewhat more efficiently than animals.
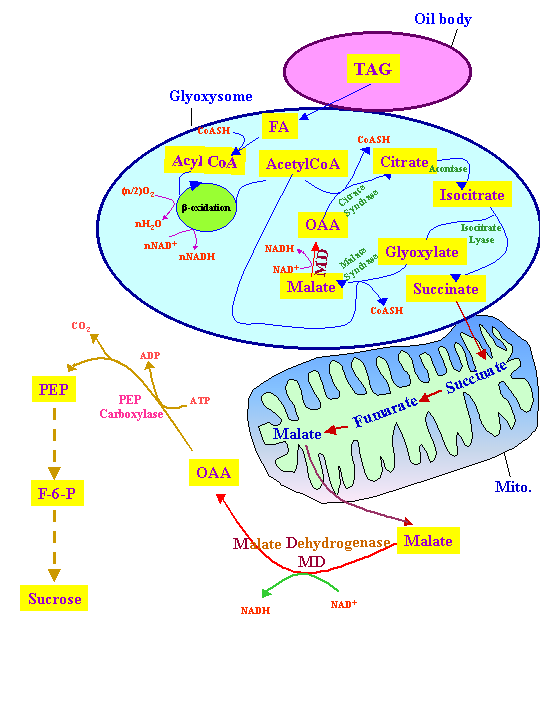
Plants, but not animals, can convert fatty acids into glucose via β-oxidation and gluconeogenesis. In this process, acetyl-CoA resulting from β-oxidation is used to convert oxaloacetate to citrate, isocitrate and succinate in glyoxysomes:
acetyl-CoA + oxaloacetate ![]() citrate
citrate ![]() isocitrate
isocitrate ![]() succinate
succinate
The glyoxylate cycle has five essential enzymes: citrate synthase (CSY), aconitase (ACO), isocitrate lyase, malate synthase, and NAD malate dehydrogenase (MDH). ACO and MDH, function in the cytosol while CSY, isocitrate lyase and malate synthase function in peroxysomes (Eckardt, 2005):
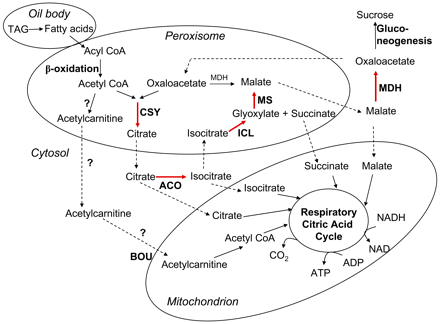
The succinate is transferred to mitochondria where it is converted to oxaloacetate (OAA). The OAA is eventually converted to triose-P and finally to glucose in the cytosol as was discussed in Respiration lectures.
Mohammed Abd-Elreheem
Unlike β-oxidation in which fatty acids are catabolized 2-C at a time, α-oxidation degrades fatty acids 1-C at a time. In mammals α-oxidation is important in the catabolism of β-methyl branched fatty acids. Although α-oxidation of fatty acids has been known to occur in plants for > 40 years, the metabolic role of this pathway is only recently beginning to be clarified. An α-dioxygenase (α-DOX) has recently been characterized from plants that catalyzes α-oxidation (Hamberg et al., 1999). This a-DOX, which is inducible by pathogens and is therefore also known as pathogen-inducible oxygenase (PIOX), catalyzes the formation of long chain aldehydes and hydroxy fatty acids as well as odd chain fatty acid catabolites (Fig. 3 Hamberg et al., 1999):
![[]](Fig3.gif)
The immediate product of this α-DOX is a dioxygenated fatty acid derivative with a hydroperoxy group on carbon 2. This is further metabolized to the products mentioned above and also can serve as a precursor for peroxygenase (see below) (Fig. 10 Hamberg et al., 1999):
![[]](Fig10.gif)
Using α-DOX transgenic Arabidopsis plants Hamberg et al. (2003) found that 1 or more α-DOX products (e.g. HOT) protect hypersensitively responding leaf tissue from undergoing excessive necrosis.
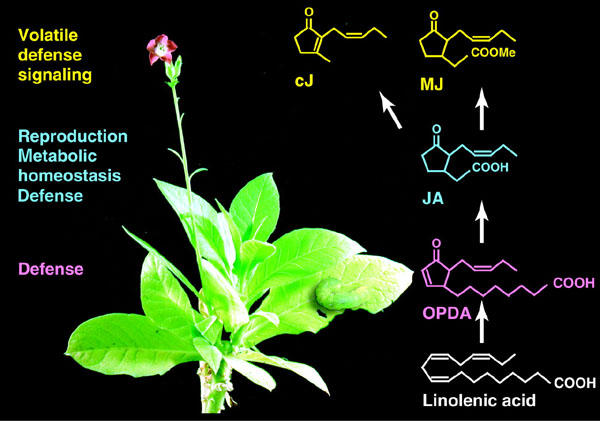
In the remainder of this lecture I will mainly focus on the formation and metabolism of additional oxylipins or oxygenated derivatives of fatty acids.
Oxylipin metabolism
Fatty acids esterified to phospholipids, especially phosphatidylcholine, can be converted by epoxygenase or hydroxylase into epoxy or hydroxy derivatives. Fatty acids can also be converted by lipoxygenase into hydroperoxy derivatives. Most lipoxygenases are thought to act on fatty acids which are first "released" by phospholipase or lipolytic acyl hydrolase before peroxidation. Lipoxygenase can peroxidize linoleic or linolenic acids forming hydroperoxy (HOO) linoleic or linolenic acids. The primary products of lipoxygenase, the fatty acid hydroperoxides usually do not accumulate in plant tissues and can be rapidly converted into additional products by one of at least four known pathways. Most plant tissues appear to have the hydroperoxide lyase and allene oxide synthase branches of the lipoxygenase pathway. To date, peroxygenases have only been demonstrated in seedlings of some plants. Hydroperoxide lyase can cleave fatty acid hydroperoxides forming six and 12 carbon aldehydes. Many plants have the capacity to convert the six carbon aldehydes into alcohols. Allene oxide synthase converts the fatty acid hydroperoxides into an unstable epoxy intermediate that can be converted into a cyclic ketone by a cyclase. A series of subsequent reactions can result in formation of jasmonic acid, which can be converted into methyl jasmonate. Peroxygenases convert fatty acid hydroperoxides into epoxides, which can be converted to hydroxides by hydroxylases. Additional enzymes that can metabolize the fatty acid hydroperoxides include divinyl ether synthase. This is illustrated in the following figure:
![[]](FA.gif)
An important aspect of fatty acid oxidation is the formation of biologically active fatty acid derivatives. This can be illustrated by the "arachidonate cascade" in animals, which generates numerous important metabolic mediators known as eicosanoids including prostaglandins (PG), thromboxanes (TX) and leukotrienes. The arachidonic cascade is initiated by release of free arachidonate by activation of phospholipase C (PLC) and diacylglycerol lipase or phospholipase A2 (PLA2) followed by oxidation by cyclooxygenase or lipoxygenase (LOX):
![[]](arach.gif)
The 1st committed step in eicosanoid formation involves oxidation of arachidonate to a hydroperoxide by LOX or cyclooxygenase.
Higher plants have a "linolenate cascade" that generally resembles the arachidonate cascade:
![[]](PMM.GIF)
LOX is an enzyme which appears to be ubiquitous in eukaryotic organisms. LOX activity has been detected in multiple tissues of all plant species examined. Cyclooxygenase has not been detected in higher plants, but 3 enzymes in plants, LOX, allene oxide synthase and allene oxide cyclase, carry out an equivalent reaction. Interestingly, the natural plant product and 1st drug, acetylsalicylic acid or aspirin, is a potent inhibitor of both cyclooxygenase in animals and the combined reactions generating prostaglandin-like products such as jasmonic acid in plants. This is also true of other analgesic agents such as acetaminophen and ibuprofen.
LOX catalyzes the peroxidation of polyunsaturated fatty acids which in higher plants and some higher fungi would be mainly linoleic (18:2) and linolenic (18:3) acids and in some cases hexadecatrienoic (16:3) acid. In animals, lower plants and lower fungi, longer chain polyunsaturated fatty acids such as arachidonic acid (20:4) (as well as 20:5 and 22:6) are more common LOX (and cyclooxygenase) substrates. The product of LOX, hydroperoxy fatty acids, are metabolized via at least four routes in higher plants to various products (see illustration below) which have biological activity and/or are important flavor and aroma compounds. These products of LOX have been referred to as octadecanoids because of their analogy to eicosanoids. Another term, oxylipins, proposed by Hamberg and Gardner, may be preferable since LOX products can be derived from 16:3. The fatty acid hydroperoxides produced by LOX can be converted into 1) keto fatty acids by other activities of LOXs; 2) hydroxy and epoxy fatty acids by cooxidative reactions of peroxygenase; 3) cleaved into two aldehydes by hydroperoxide lyase (HL) or 4) converted into an unstable allene oxide by allene oxide synthase (AOS). The biological activities of the keto fatty acids are unknown. The hydroxy and epoxy fatty acids produced by peroxygenase may have a role in cutin biogenesis and pest defense. HL products have a role in pest defense and appear to act as pollinator and herbivore attractants especially for flowers and fruits. They are also important flavor and aroma compounds in foods, beverages and perfumes. Products of AOS, especially jasmonic acid (JA) and methyl jasmonate (MJ), have generated intense interest and appear to have numerous biological activities. These include:
Oxylipin Formation from Linolenic Acid in Plant Tissues [IF: Isomerization factor(s)]:
Click on this link for a larger,clearer image: OxylP
![[]](oxylp.gif)
The mechanism of the LOX (lipoxygenase) catalyzed polyunsaturated fatty acid peroxidation is fairly well understood. LOX is a non-heme single iron containing dioxygenase which catalyzes the peroxidation of cis,cis -1,4 pentadiene structures:
In its oxidized state, Fe3+, LOX can abstract a
proton from a PUFA such as linolenic acid. In the following example,
a 13-LOX abstracts a proton from C11 of linolenic acid reducing LOX (![]() Fe2+) and producing a fatty acid free
radical. While the substrate is still bound to LOX, the free radical
shifts to C13 and the 12-13 cis double bound shifts to C11-C12 and
becomes trans forming a conjugated diene structure. In the
presence of O2 a peroxy radical can be formed that can be
reduced to a hydroperoxide by the bound LOX enzyme. This results in the
reoxidization of LOX which is then released.
The hydroperoxy fatty acid can be cleaved into two aldehydes by
hydroperoxide lyase also common in plant tissues:
Fe2+) and producing a fatty acid free
radical. While the substrate is still bound to LOX, the free radical
shifts to C13 and the 12-13 cis double bound shifts to C11-C12 and
becomes trans forming a conjugated diene structure. In the
presence of O2 a peroxy radical can be formed that can be
reduced to a hydroperoxide by the bound LOX enzyme. This results in the
reoxidization of LOX which is then released.
The hydroperoxy fatty acid can be cleaved into two aldehydes by
hydroperoxide lyase also common in plant tissues:
![[]](glycerolipids.gif)
Of particular interest recently in plant biology has been the AOS or jasmonic acid pathway. This is increasingly being recognized as being of fundamental importance in signal transduction in response to biotic and abiotic stresses. The formation of these compounds is frequently elicited by pest attack including wounding and other stresses causing transient accumulation of intermediates of the "linolenate cascade" (see section 17.8 of the class text):
Especially interesting is the induced release of phermone-like compounds that attract predators and parasites to plants being fed upon by herbivorous insects. Koch et al. (1999) find that there are 4 groups of structurally distinct types of compounds from the linolenate cascade with different effects on release of attractants to beneficial insects:
a.) 18:3 and acyclic derivatives.
b.) C18 cyclopentanoid fatty acids.
c.) C12 cyclopentanoid fatty acids especially epi-JA and JA.
d.) Amino acid conjugates of linolenate cascade intermediates.
This is illustrated in Fig. 1 of Koch et al. (1999):
![[]](Fig1a.jpg)
Do you see anything wrong with this figure?
Reading Assignment for the 1st nitrogen metabolism lecture:
a) REQUIRED:
1 - Hu, Y., A.W. Fay, C.C. Lee, J. Yoshizawa, and M.W. Ribbe. 2008. Assembly of Nitrogenase MoFe Protein. Biochemistry 47:3973-3981..
2 - Chapter 16, sections 16.1 - 16.5 of the Biochemistry & Molecular Biology of Plants class text.
b) SUGGESTED:
1 - Tezcan, F.A., J.T. Kaiser, D. Mustafi, M.Y. Walton, J.B. Howard, and D.C. Rees. 2005. Nitrogenase Complexes: Multiple Docking Sites for a Nucleotide Switch Protein. Science 309: 1377-1380.
2 - Schrock, R.R. 2006. Nitrogen Fixation Special Feature: Reduction of dinitrogen. PNAS 103:17087.
3 - Cullimore, J. and J. Denarie. 2003. How legumes select their sweet talking symbionts. Science 302:575-578.
4 - Szczyglowski, K. and L. Amyot. 2003. Symbiosis, Inventiveness by Recruitment? Plant Physiol. 131: 935-940 .
5 - Weare, W.W., X. Dai, M.J. Byrnes, J.M. Chin, R.R. Schrock, and P. Muller. 2006. Nitrogen Fixation Special Feature: Catalytic reduction of dinitrogen to ammonia at a single molybdenum center. PNAS 103:17099-1710.
6 - Hipkin, C.R., D.J. Simpson, S.J. Wainwright, and M.A. Salem. 2004. Nitrification by plants that also fix nitrogen. Nature 430: 98-101.
7 - Seefeldt, L.C., I.G. Dance and D.R. Dean. 2004. Substrate Interactions with Nitrogenase: Fe versus Mo. BIOCHEMISTRY 43: 1401-1409.
8 - Mylona, Panagiota, Katharina Pawlowski and Ton Bisseling. 1995. Symbiotic Nitrogen Fixation. The Plant Cell 7, pp. 869-885.
| All materials © 2013 David Hildebrand, unless otherwise noted. | |||||||
| home | syllabus | lecture schedule & web notes | virtual office hours | messages & answers from the instructor | supplementary material | related links | What's new? |
{kind=link}