
Lecture Twenty-four
Nitrogen Metabolism -- Essential and non-protein
amino acid biosynthesis; other N compounds
Goal: Briefly review the common protein amino acids and their properties. To gain a good working knowledge and understanding of the biosynthesis of essential amino acids and other selected important nitrogenous compounds of plant tissues.
Outline:
Background Readings for the discussion on amino acid and other N compound metabolism :
a) REQUIRED (not required in 2016):
1 - Chapter 8 and chapter 9, section 9.5 of the Biochemistry & Molecular Biology of Plants class text.
b) SUGGESTED:
1 - Smart, D.R. and A. J. Bloom. 2001. Wheat leaves emit nitrous oxide during nitrate assimilation. PNAS 98: 7875-7878.
2 - Lam et al. 1995. Use of Arabidopsis mutants and genes to study amide amino acid biosynthesis. The Plant Cell 7, pp. 887-898.
3 - Guo, F.Q., M. Okamoto and N.M. Crawford. 2003. Identification of a plant Nitric oxide synthase gene involved in hormone signaling. Science 302: 100-103.
4 - Shewry et al. 1995. Seed storage proteins: structures and biosynthesis. The Plant Cell 7, pp. 945-956.
5 - von Wettstein et al. 1995. Chlorophyll biosynthesis. The Plant Cell 7, pp. 1039-1057.
6 - Zeier, J., M. Delledonne, T. Mishina, E. Severi, M. Sonoda, and C. Lamb. 2004. Genetic Elucidation of Nitric Oxide Signaling in Incompatible Plant-Pathogen Interactions. Plant Physiol. 136: 2875-2886.
7 - Jasid, S., M. Simontacchi, C.G. Bartoli, and S. Puntarulo. 2006. Chloroplasts as a Nitric Oxide Cellular Source. Effect of Reactive Nitrogen Species on Chloroplastic Lipids and Proteins. Plant Physiol. 142:1246-1255.
8 - Peng Pan, Eilika Woehl and Michael F. Dunn. 1997. Protein architecture, dynamics and allostery in tryptophan synthase channeling. TIBS 22: 22-27.
Remember from lectures 22 and 23, N enters roots as NO3- or NH4+. The NH4+ is incorporated into amino acids in roots and leaves and the amino acids accumulate in proteins. The main if not sole function of some proteins is to provide a store of amino acids (see below).
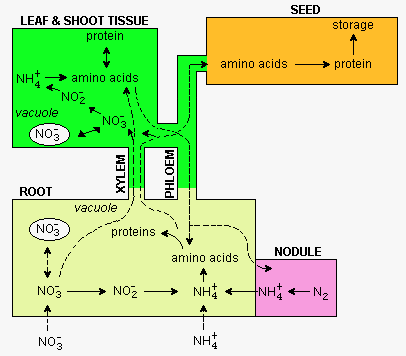
As mentioned in lectures 22 and 23, NH4+ formed as a result of nitrate reduction or N2 fixation is 1st incorporated into amino acids via the glutamine synthetase (GS) reaction:
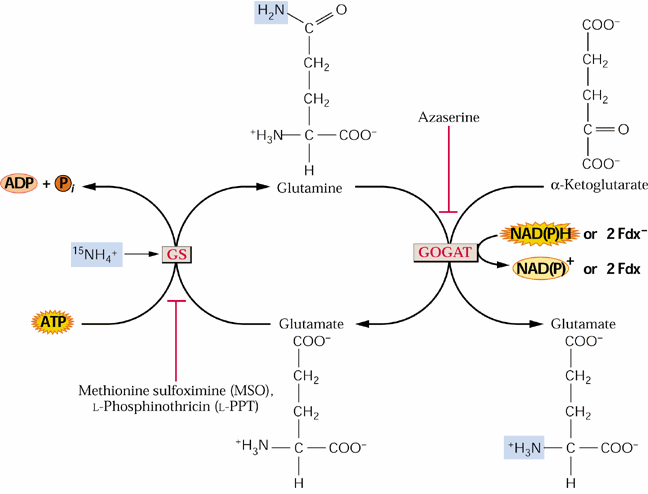
The glutamate consumed by the GS reaction is replenished by an alternative mode of glutamate synthesis GOGAT glutamate: oxoglutarate aminotransferase).
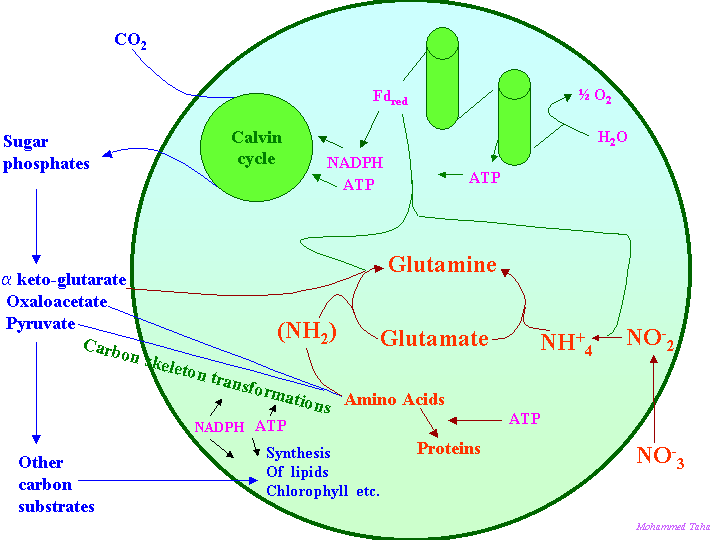
A general scheme of amino metabolism in relation to general metabolism in plant cells is given in the following figure:
Plants contain multiple isoenzymes of GS, some located in the cytoplasm (=GS1) and another in plastids (= GS2). All plants examined contain at least 3 GS1 isozymes, but only one nuclear encoded GS2 isoenzyme.
]
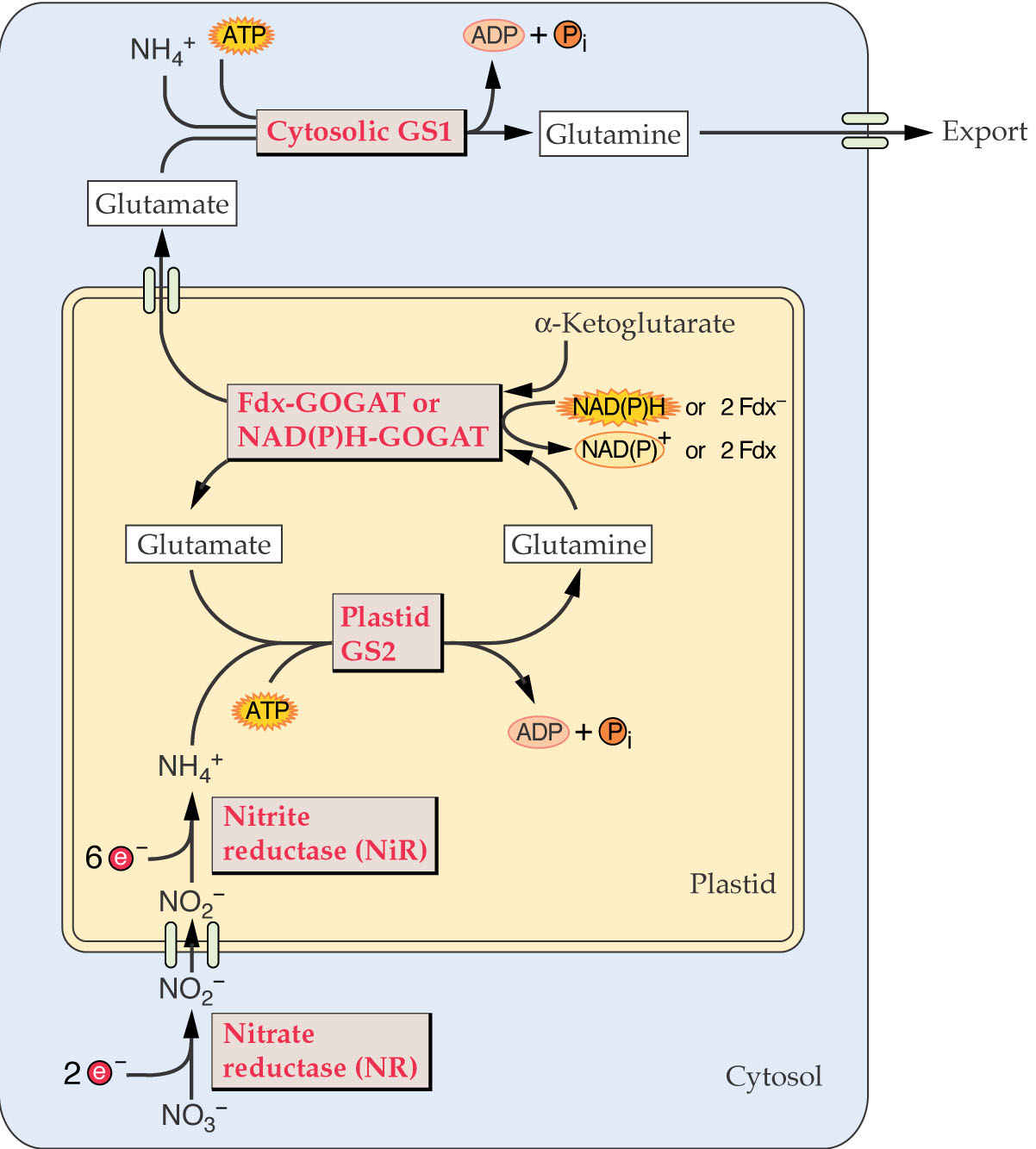
GS2 appears to be most important in ammonia assimilation, but GS1 isozymes are also functional. In C3 plants ammonium production from photorespiration can exceed that formed by nitrite reduction by 10-fold! Where is the ammonium formed in photorespiration? Could the ammonium be assimilated by other enzymes?
The GS2 of plastids has been shown to also be responsible for photorespiratory ammonia assimilation in leaves by analysis of GS2 mutants. How can these be grown? GS1 in leaves is not expressed in mesophyll cells and apparently is only expressed in phloem cells of leaves.
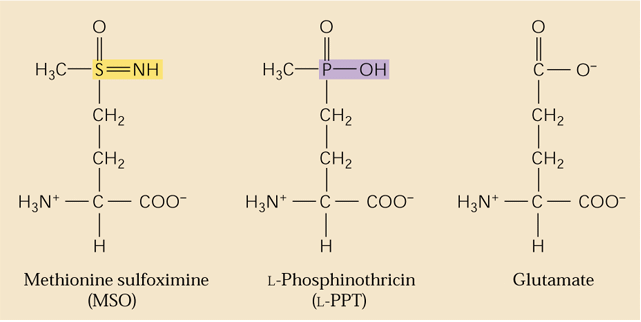
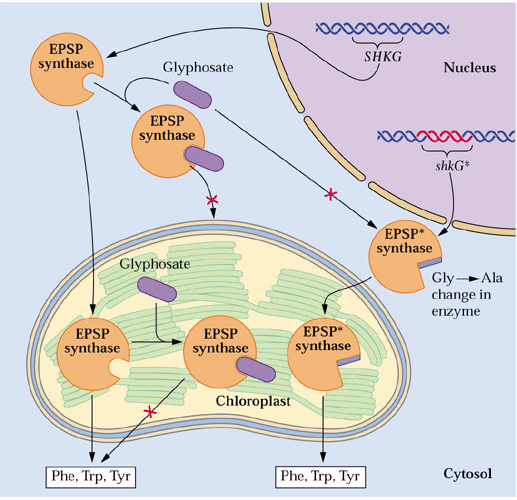
GS is the target of a major herbicide Basta® (AgrEvo Co.). The active ingredient in Basta®, Rely®, Finale®, Liberty® and Ignite® is glufosinate-ammonium. Glufosinate (= phosphinothricin) is a substrate analogue of glutamate and strong inhibitor of GS causing toxic levels of ammonium to buildup in treated plants. This can be a natural product produced by Streptomyces ( Lee et al., 2009). Thus it is a broad-spectrum herbicide. Glufosinate is rapidly degraded in the soil leaving behind no toxic residues. Crops such as corn and canola are now being marketed that have been genetically engineered for phosphinotricine or Basta resistance. Such corn hybrids are being marketed under the trade name Liberty-Link® corn. Methionine sulfoximine also inhibits GS and azaserine inhibits GOGAT (see above). Would phosphinothricin be expected to affect metabolism in animals including humans? Name some other amino acid biosynthetic enzymes that are targets of herbicides
]
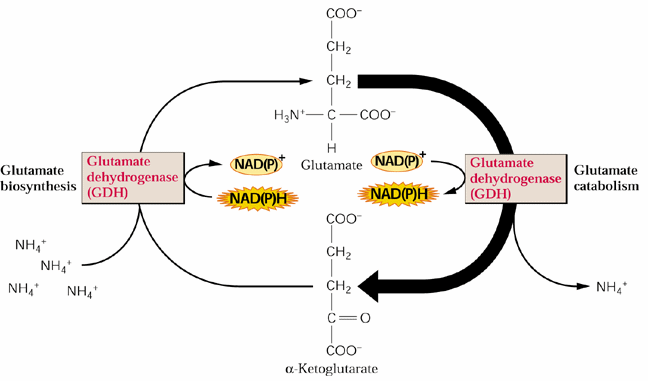
GDH is thought to be primilarly catabolize (deaminate) glutamate. Why?
Proteins commonly contain 20 different amino acids. Of these 20, 10 (termed essential amino acids) cannot be synthesized by mammals and must be obtained from the diet. The discussions on amino acid biosynthesis will emphasize essential amino acid biosynthesis in plants.
See Fig. 8.1 of the class text (higher res). Which amino acids are "essential"?
All amino acids contain a basic amino group and an acid carboxylic acid group and have the general formula:
The R substituent has a wide range of chemical properties ranging from H in glycine, the simplest amino acid, to the indole group of tryptophan.
Amino acids are placed into four groupings based on physical/chemical properties:
Proteins are polypeptides composed of > ~ 20-30 amino acids joined by peptide bonds. Peptide bonds are formed in dehydration reactions similar to the glycosidic bond of polysaccharides in which an amide bond is formed between the carboxyl group of one amino acid and the amino group of another:
The amino acid composition of proteins determines its physical properties in three important ways.
1. The size or molecular weight of a protein
depends on the number (and size) of its amino acids.
2. The charge or isoelectric point, pI, depends on the relative proportion
of basic and acidic amino acids.
3. The hydrophobicity is determined by the relative levels of hydrophobic and hydrophilic amino acids.
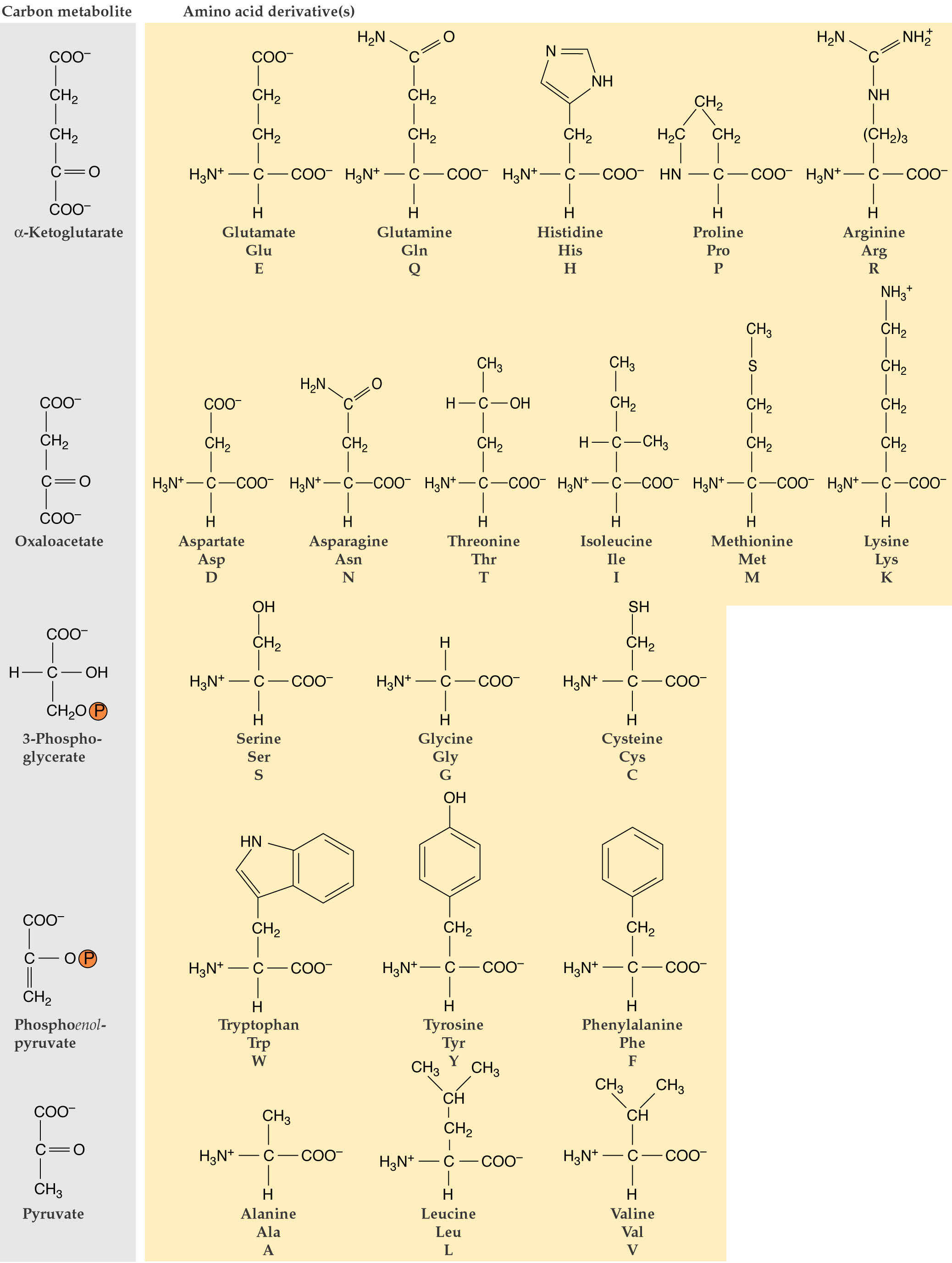
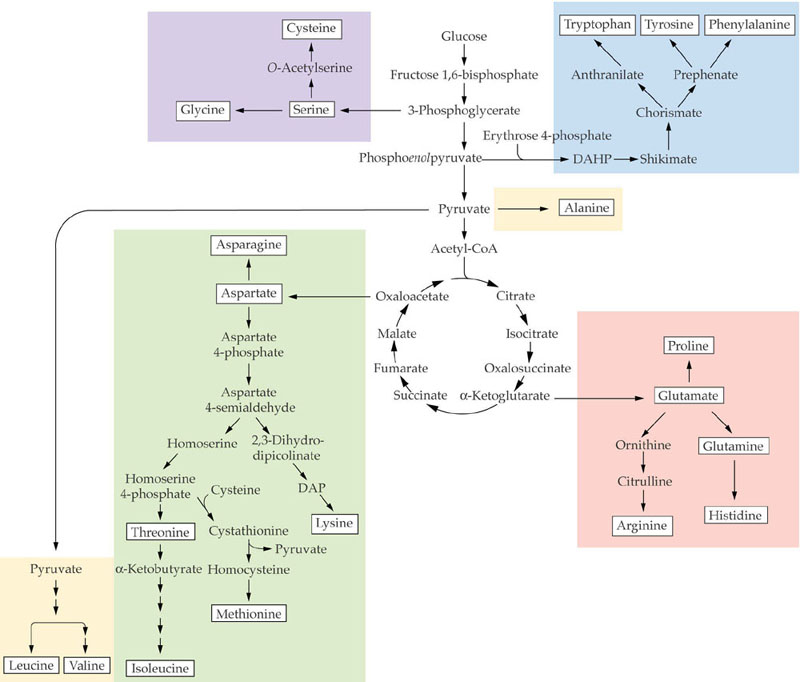
Amino acids can be placed into five families based on biosynthetic origins (see Fig. 8.2):
Histidine does not fall within any of these families as it has no other amino acid as an immediate biosynthetic precursor.
Essential Amino Acid Biosynthesis
The carbon skeletons for the various amino acids come directly or indirectly from intermediates of the Calvin-Benson cycle. Of these, 3-phosphoglycerate is the most important carbon precursor in amino acid biosynthesis providing carbon skeletons for 16 of the 20 amino acids. The 3-phosphoglycerate produced by the Calvin cycle can be exported from plastids by a triose-phosphate translocator located in the plastid envelope. It can be converted into phosphoenolpyruvate (PEP) in the cytosol.
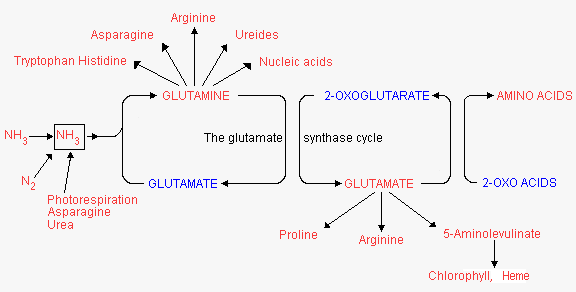
From PEP there are 2 pathway branches one leading to pyruvate and the other to oxaloacetate.
Additionally, PEP + erythrose 4-phosphate are precursors of the synthesis of aromatic amino acids via the shikimate pathway. The oxaloacetate has 2 functions in amino acid synthesis. 1) It is transaminated to aspartate, the precursor of the aspartate family amino acids; and 2) together with pyruvate it is the precursor of a-KG which can be transaminated to glutamate as described in Lecture 21. Glutamate is the precursor of the glutamate family amino acids. Phosphoglycolate is the precursor of the serine family amino acids and ribose 5-phosphate for histidine.
Amino acid origins:
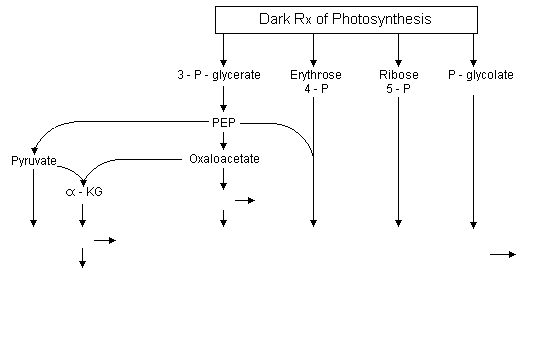
|
|
||||||||||||||||||||||
These intermediates of amino acid biosynthesis involve multiple subcellular sites. The pyruvate and oxaloacetate are formed in the cytoplasm from 3-phosphoglycerate originally made in chloroplasts by the Calvin cycle. Some pyruvate is translocated into mitochondria where it is converted to acetyl-CoA which can condense with oxaloacetate, forming citrate. Citrate is transferred to the cytoplasm in exchange for oxaloacetate where it is converted into a-KG. Citrate from the citric acid cycle can also be converted into a-KG in mitochondria:
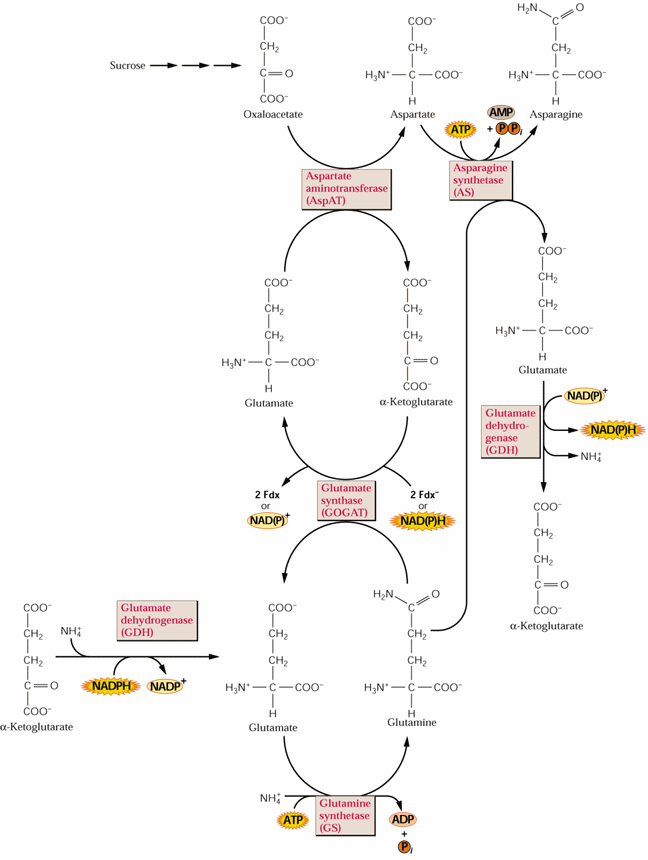
Glutamate formed from glutamine and α-KG, as described in Lecture 23, is a precursor for the synthesis of proline, arginine and aspartate. Aspartate is synthesized from glutamate and oxaloacetate by an aminotransferase reaction:
Glutamate + oxaloacetate ![]() α-KG + aspartate
α-KG + aspartate
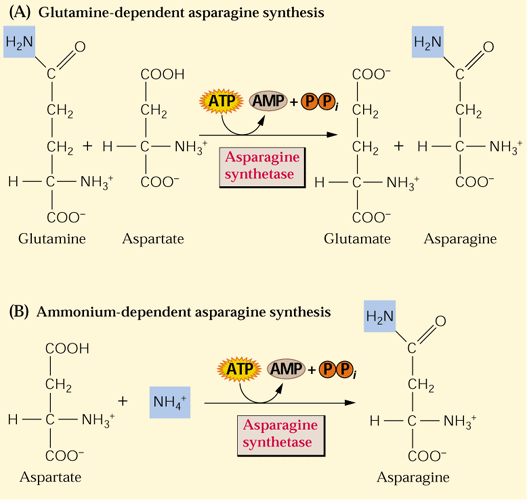
Asparagine synthesis simply involves the transfer of another amino group from glutamine to aspartate in an ATP-dependent reaction (see Fig. 8.4 above). Asparagine synthesis increases in the dark:
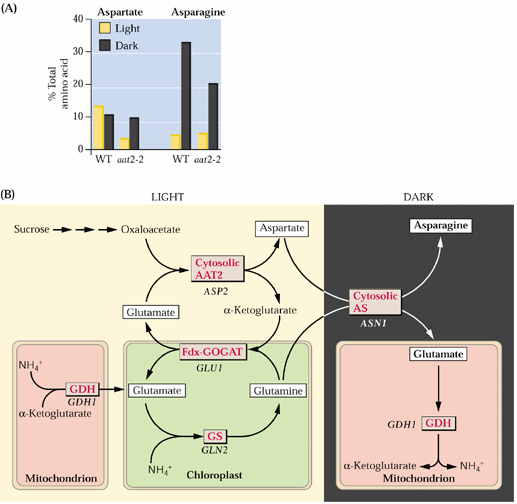

Asparagine can be synthesized directly from ammonium but preferentially utilizes Q (gln) as the N source:
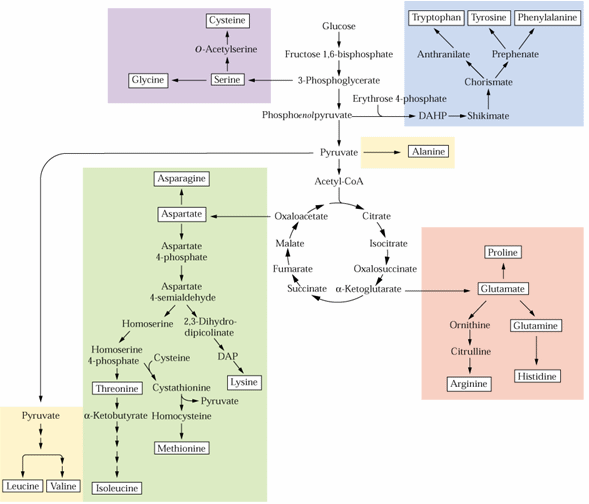
Aspartate is a precursor for the synthesis of 4 additional amino acids in plastids methionine, lysine, threonine and isoleucine (see Galili 1995. The Plant Cell 7:899-906 and following figure):
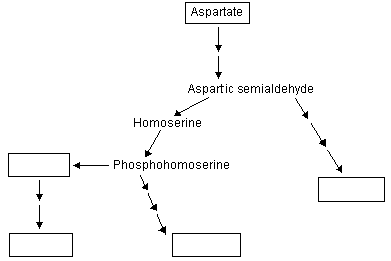
|
|
||||||
The 1st step involves aspartate kinase and this and certain subsequent enzymes in the biosynthesis of threonine and lysine are feedback inhibited by these 2 amino acids:
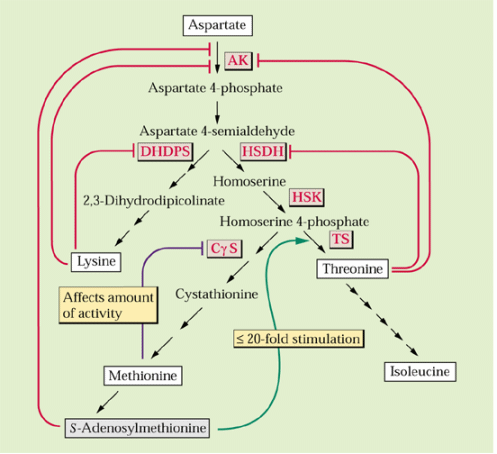
]
The 3 aromatic amino acids, phenylalanine, tyrosine and tryptophan, are synthesized from the shikimate pathway which is only found in microorganisms and plants. The shikimate pathway is not only a source of these amino acids also of many phenolic compounds including pigments, defensive compounds and especially lignin. Under normal growth conditions ~20% of the carbon fixed by plants flows through this pathway. The shikimate pathway involves 7 enzymatic steps apparently located in plastids (Figure 1 of Herrmann 1995. The Plant Cell 7:907-919). This pathway which leads to phenylpropanoids was described in lectures 20 and 21 and generally by Guillet et al. (2000).
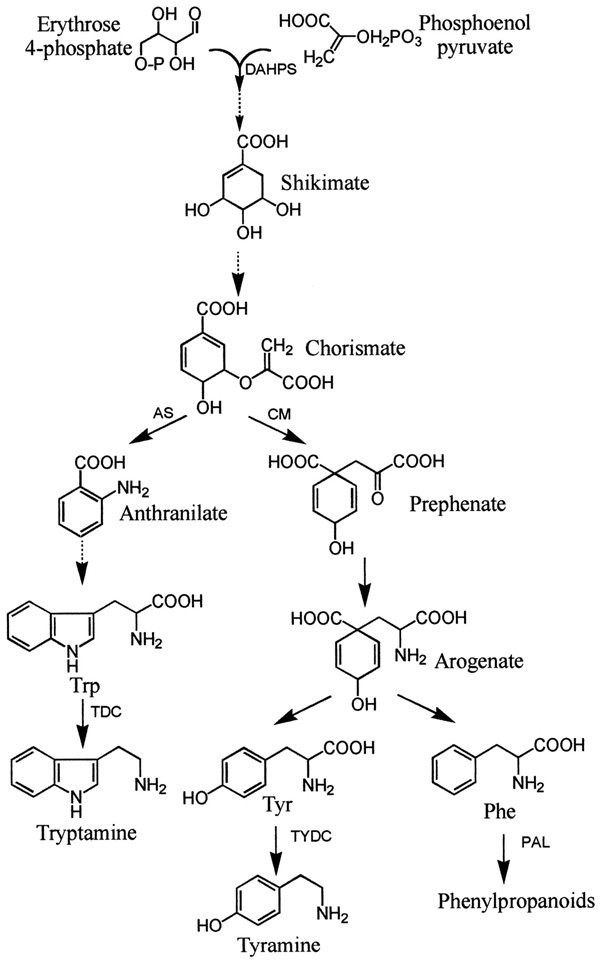
Aromatic amino acid biosynthesis
The penultimate step of chorismate biosynthesis is catalyzed by 5-enolpyruvylshikimate-3-P synthase (EPSPS)
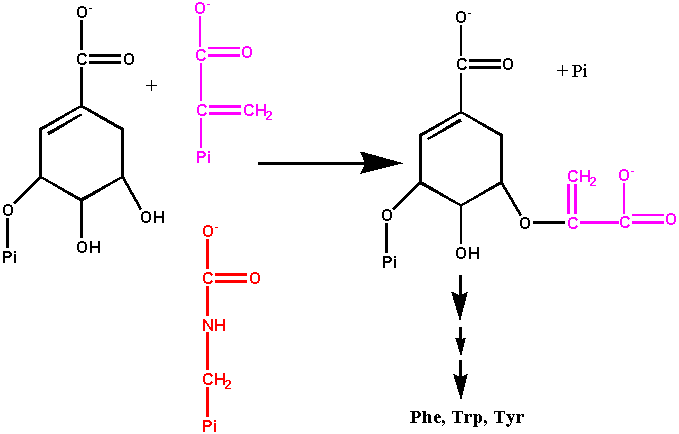
]
Dicamba resistant crops...
Tryptophan biosynthesis is perhaps the best understood amino acid biosynthetic pathway in plants (see section 8.3 of the class text). The synthesis from chorismate involves 6 plastid localized enzymatic steps as illustrated in Figure 2 of Radwanski and Last (1995, The Plant Cell 7:921-934) and Fig. 8.33 of the class text. The 1st step involves addition of an amine and the third ring double bound forming anthranilate. The rest of the indole ring structure construction starts with addition of phosphoribose. In the 3rd step the phosphoribosyl group is rearranged followed by closure of the indole ring in step 4. The last 2 steps are catalyzed by subunits of tryptophan synthase with the 5th step involving removal of glycerol-3-P group forming indole and the 6th the addition of serine to the indole ring producing tryptophan. The indolic phytoalexins are formed from Trp.
Tryptophan biosynthesis in enteric bacteria represents a classical example of true biochemical "channeling". As Pan et al. (1997) explain:
"The α2β2 form of the tryptophan synthase bienzyme complex catalyses the last two steps in the synthesis of L-tryptophan, consecutive processes that depend on the channeling of the common metabolite, indole, between the sites of the α- and β-subunits through a 25 Å-long tunnel. The channeling of indole and the coupling of the activities of the two sites are controlled by allosteric signals derived from covalent transformations at the β-site that switch the enzyme between an open, low-activity state, to which ligands bind, and a closed, high-activity state, which prevents the escape of indole."
Storage Protein Catabolism
Plants usually store amino acids in proteins in storage tissues. Also plants often recycle amino acids from turnover of proteins in senescing tissues such as leaves. Plants contain 5 major classes of endoproteinases (reviewed by Callis 1995, The Plant Cell 7:921-934 and in section 9.5 of the class text):
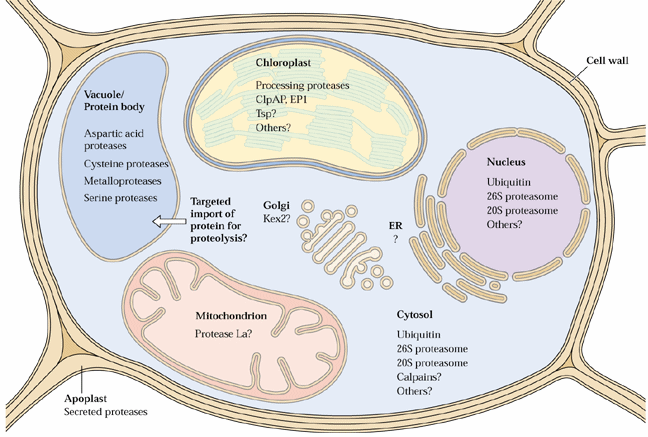
The proteosome proteolytic complexes are known in nuclei and the cytosol. It is not yet known if they also can be found in chloroplasts, mitochondria or ER.
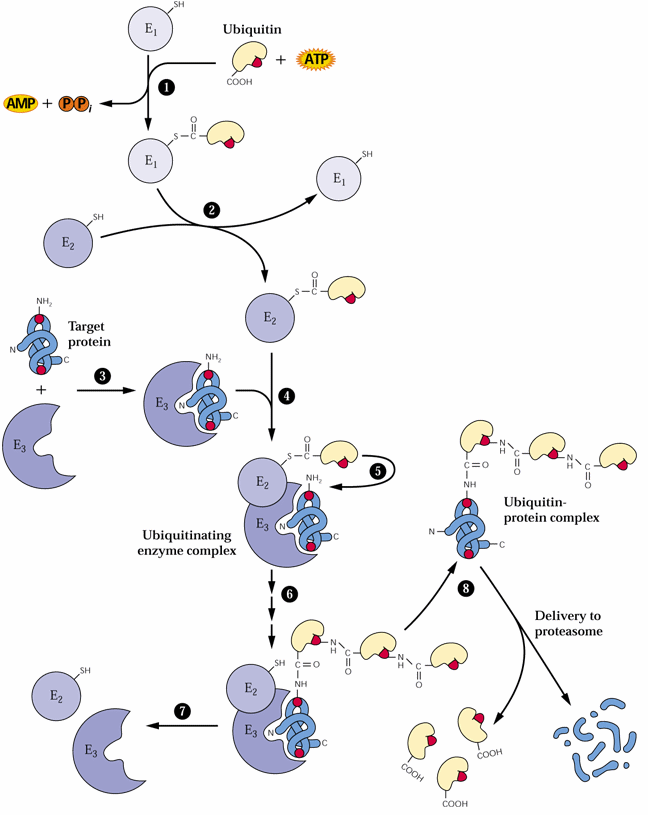
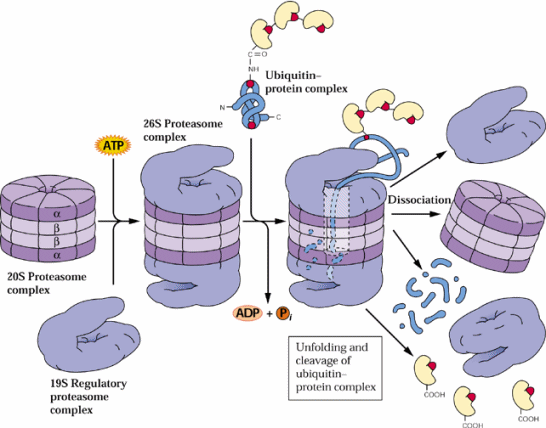
These endoproteinases often cause considerable problems in purification and biochemical characterization of plant proteins. Proteinase inhibitors are available to many of these endoproteinases. Serine proteinase inhibitors include phenylmethylsulfonyl fluoride (PMSF), trypsin inhibitors, aprotinin and leupeptin. Cysteine proteinases can be inhibited by iodoacetate and iodoacetamide and aspartic acid proteinase by pepstatin. Metallo-proteinase inhibitors include 1,10-phenanthroline and EDTA. Recently a number of biochemical suppliers have introduced handy (and rather expensive) proteinase inhibitor cocktails to facilitate protein biochemistry studies since many tissues have a range of proteinases which need an array of proteinase inhibitors.
Protein degradation is an important factor in plant development as reported by Hellmann and Estelle in their 2002 Science article.
Non-Protein Amino Acids
The proteins of all organisms are based on the 20 common L-amino acids although some can be modified post-translationally. However some plants, especially members of the Leguminosae and Liliaceae, make many different amino acids that are not incorporated into proteins. These so-called non-protein amino acids can accumulate to high levels in storage tissues such as seeds and rhizomes at up to 10% of the dry weight and 50% of the nitrogen. Two examples of non-protein amino acids are 3-cyanoalanine and L-canavanine . Note the structural resemblance of 3-cyanoalanine to L-asparagine:
And L-canavanine to L-arginine:
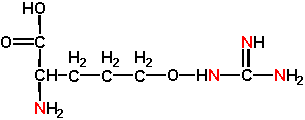
These non-protein amino acids mainly have roles in plant defense, but can also act as reduced N storage structures. They have multiple anti-pest actions as covered in the text including inhibition of aminoacyl-tRNA synthetases or other steps of protein synthesis. In other cases they can be incorporated into proteins in place of normal amino acids resulting in defective proteins. Plants that synthesize these non-protein amino acids manage to circumvent any toxicity of these compounds by having amino acyl-tRNA synthetases which neither are inhibited by non-protein amino acids nor incorporate them into proteins. Some insects have evolved similar adaptations such as a bruchid beetle and a weevil that can feed on seeds of the legume, Dioclea megacarpa, with levels of canavanine toxic to other insects (Rosenthal, UK, Dept. Biology).
Amines
Amines can be classified as aliphatic mono- and diamines, polyamines, aromatic amines and amine conjugates:
Amines are cationic at physiological pHs having pKa values of 9-11. Aliphatic monoamines are especially abundant in flowers of certain plants such as in the Araceae and Rosaceae. They have an odor of rotting flesh and are attractive to certain creatures that serve as pollinators. A principal biosynthetic route in higher plants involves transfer of an amino group from alanine to an aldehyde in mitochondria producing an amine and keto acid:
The polyamines spermidine and spermine appear to be ubiquitous in plants as does the diamine, putrescine. The diamine, cadaverine, has only been found in legumes:
Would you expect these molecules to have differences in odors?
What might you expect these molecules to do in cells?
In bacteria putrescine and polyamines play a role in DNA stabilization and chromosome structure, as do histones in nuclear chromosomes. Chloroplasts and mitochondria do not have histones and putrescine and polyamines may serve this role in these organelles. Polyamines can also stimulate protein synthesis likely by interactions with RNA. L-Arginine can be a precursor of putrescine, and L-lysine or L-homoarginine of cadaverine. The condensation of S-methyl-S-adenosyl homocysteamine (SMAH) [from decarboxylation of S-adenosyl-L-methionine (SAM) (see Lect. 25)] with putrescine forms spermidine and the condensation of spermidine with SMAH forms spermine. Putrescine (and spermidine and spermine) can also be derived from decarboxylation of the non-protein amino acid, ornithine, as will be described in Lecture 26.
Aromatic amines are usually produced by decarboxylation of amino acids. For example dopamine, tryptamine and histamine are formed from 3,4-dihydroxyphenylalanine, tryptophan or histidine:
Histamine and serotonin, formed by hydroxylation of tryptamine, are important neurotransmitters. The "sting" of stinging nettles (Urtica dioica and other spp.) is derived from accumulation of histamine, serotonin and acetylcholine in bulbs or glands at the ends of hairs or trichomes on leaves. When one brushes by these plants, these neurotransmitters are injected into the skin immediately causing painful itching and swelling.
In Lecture 11 we discussed release of the toxic cyanide from cyanogenic glucosides also known as cyanogens. Cyanogenic glucosides are synthesized from amino acids and monosaccharides in microsomes:
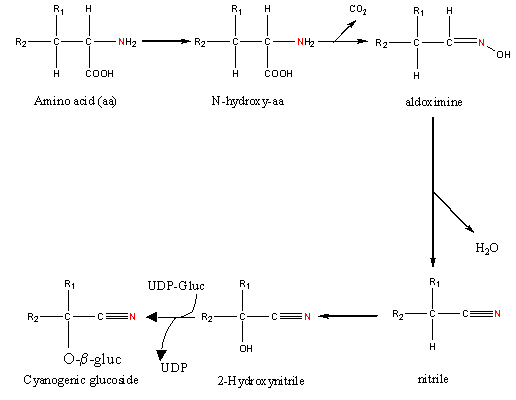
This involves N-hydroxylation of an amino acid followed by decarboxylation forming an aldoximine. The aldoximine is dehydrated to a nitrile that is then hydroxylated. A cyanogenic glycoside is formed between the hydroxy group of 2-hydroxynitrile and the monosaccharide of a UDP-glucoside.
Heme
The porphyrin heme is another important N compound. The synthesis of heme is the same as that for chlorophyll (described in Lecture 2 and by von Wettstein et al. 1995 of the Plant Cell) up to protoporphyrin IX. Protoporphyrin IX is converted to heme by addition of the iron by ferro-chelatase. Heme is an essential component of many oxygen involving proteins such as cytochromes.
Chlorophylls are Mg+2 containing substituted tetrapyrroles structurally related to hemes, Fe+2 containing porphyrins. The structure of chlorophyll is given in Fig. 1 of the article by von Wettstien et al. (1995) of the Plant Cell:
![[Image of the chlorophyll molecule]](fig1.jpg)
Chlorophyll is synthesized in chloroplasts from 8 molecules of 5-aminolevulinic acid. The 8 red lines indicate the location of the parts of 5-aminolevulinic acid in the finished molecule. Position 3 that is a methyl group in chlorophyll a (illustrated) is a formyl (CHO) group in chlorophyll b. In higher plants 5-aminolevulinic acid is synthesized from glutamic acid as illustrated in Fig. 4 of von Wettstien et al. (1995):
![[Image
of 5-aminolevulinic acid synthesis]](fig4.jpg)
Nitric and Nitrous Oxide
Nitric oxide (NO) is a recently discovered signaling molecule in animals. NO is involved in both physiological and pathophysiological conditions including inflammation, acute responses and programmed cell death. NO is among the very interesting group of compounds known as reactive oxygen species (ROS). ROS appear to act as diffusible intercellular signals for activation of various defense genes in animals, bacteria and plants. Can you name a ROS that recently has been proposed to activate defense responses in plants?
NO has unique chemistry that allows both stability and reactivity at the same time. Because of this, NO and its redox-activated forms, have been regarded as the only ROS that can fulfill the requirements of a true intra- and inter- cellular signaling molecule.
NO can be formed two ways in plants:
1. NO can be produced nonenzymatically through light-mediated conversion of NO2 by carotenoids or
2. NO can be formed enzymatically from NO2 by NADPH nitrate reductases.
Evidence has been accumulating implicating NO as an effector in plant growth, development and defense. Durner, Wendehenne and Klessig at Rutgers recently demonstrated defense gene induction in tobacco by NO (Jörg Durner, David Wendehenne, and Daniel F. Klessig. 1998. Defense gene induction in tobacco by nitric oxide, cyclic GMP, and cyclic ADP-ribose PNAS 95: 10328-10333).
What is it about the structure of NO that gives rise to its unique properties?
Last assignment for 2004: Re-read the Guo et al. (2003) paper (citation above) if needed and describe the reaction catalyzed by ATNOS1 according to Guo et al. (2003). Give the structures of the substrates and products. Briefly what are the phenotypic consequences of Arabidopsis plants unable to carry out this reaction? (5 points)
Nitrous oxide (N2O) is a key
atmospheric greenhouse gas that contributes to global climatic change through
radiative warming and depletion of stratospheric ozone. Smart
and Bloom (2001) report a correlation of leaf N2O
emissions with leaf nitrate
assimilation activity, as measured by using the assimilation
quotient, the ratio of CO2 assimilated to O2 evolved.
In vitro production of N2O by both intact chloroplasts and
nitrite reductase, but not by nitrate
reductase, indicated that N2O produced by leaves occurred
during photoassimilation of NO![]()
![]()
![]()
Background Readings for the discussion on N/S compounds :
a) REQUIRED:
1 - Chapter 16 sections 16.10 - 16.14 of the Biochemistry & Molecular Biology of Plants class text.
b) SUGGESTED:
1 - Stephane Ravenel, Bertrand Gakiere, Dominique Job and Roland Douce. 1998. The specific features of methionine biosynthesis and metabolism in plants. PNAS 95: 7805-7812.
2 - Thomas Leustek, Melinda N. Martin, Julie-Ann Bick and John P. Davies. 2000. PATHWAYS AND REGULATION OF SULFUR METABOLISM REVEALED THROUGH MOLECULAR AND GENETIC STUDIES. Annu. Rev. Plant Physiol. Plant. Mol. Biol. 51: 141-165.
3 - Naoko Yoshimoto, Eri Inoue, Kazuki Saito, Tomoyuki Yamaya and Hideki Takahashi. 2003. Phloem-Localizing Sulfate Transporter, Sultr1;3, Mediates Re-Distribution of Sulfur from Source to Sink Organs in Arabidopsis1. Plant Physiol. 131: 1511-1517.
4 - Kopriva, S., M. Suter, P. von Ballmoos, H. Hesse, U. Krahenbuhl, H. Rennenberg, and C. Brunold. 2002. Interaction of Sulfate Assimilation with Carbon and Nitrogen Metabolism in Lemna minor. Plant Physiology. 130: 1406-1413.
5 - Anna Koprivova, Marianne Suter, Roel Op den Camp, Christian Brunold, Stanislav Kopriva. 2000. Regulation of Sulfate Assimilation by Nitrogen in Arabidopsis. Plant Physiol. 122: 737-746.
6 - Shibagaki, N., A. Rose, J.P. McDermott, T. Fujiwara, H. Hayashi, T. Yoneyama and J.P. Davies. 2002. Selenate-resistant mutants of Arabidopsis thaliana identify Sultr1;2, a sulfate transporter required for efficient transport of sulfate into roots. Plant J. 29: 475-486.
| All materials © 2016 David Hildebrand, unless otherwise noted. | |||||||
| home | syllabus | lecture schedule & web notes | virtual office hours | messages & answers from the instructor | supplementary material | related links | What's new? |
{kind=link}
{kind=link}