
Lecture Twenty
Synthesis of Membrane Lipids and Triglycerides
The purpose of today's lecture/discussion is to cover important aspects of the biosynthesis of major plant membrane lipids and storage oil. An alternative pathway for saturated fatty acid synthesis in plants will be briefly discussed as an example of the determination of such biosynthetic processes.
__________________________________________________________________________________________________________________
Reading Assignments for Lecture 20 discussion:
a) REQUIRED:
1 - Bates, P.D., Fatihi, A., Snapp, A.R., Carlsson, A.S., Browse, J., and Lu, C. (2012). Acyl Editing and Headgroup Exchange Are the Major Mechanisms That Direct Polyunsaturated Fatty Acid Flux into Triacylglycerols. Plant Physiology 160, 1530-1539.
2 - Chapter 8, sect 8.5, 8.7 & 8.8 (Chapter 10, sections 10.5 - 10.8 and 10.10.1 -10.10.3 2000 text), of the Biochemistry & Molecular Biology of Plants class text.
b) OPTIONAL:
1 - Lager, I., Yilmaz, J.L., Zhou, X.-R., Jasieniecka, K., Kazachkov, M., Wang, P., Zou, J., Weselake, R., Smith, M.A., Bayon, S., et al. (2013). Plant Acyl-CoA:Lysophosphatidylcholine Acyltransferases (LPCATs) Have Different Specificities in Their Forward and Reverse Reactions. Journal of Biological Chemistry 288, 36902-36914.
2 - Zhang, M., J. Fan, D.C. Taylor, and J.B. Ohlrogge. 2009. DGAT1 and PDAT1 Acyltransferases Have Overlapping Functions in Arabidopsis Triacylglycerol Biosynthesis and Are Essential for Normal Pollen and Seed Development. Plant Cell 21:3885-3901.
3 - Lu, C., Z. Xin, Z. Ren, M. Miquel, and J. Browse. 2009. An enzyme regulating triacylglycerol composition is encoded by the ROD1 gene of Arabidopsis. Proceedings of the National Academy of Sciences 106:18837-18842.
4 - Engelman, D.M. 2005. Membranes are more mosaic than fluid. Nature 438: 578-580.
5 - McMahon, H.T., and J.L. Gallop. 2005. Membrane curvature and mechanisms of dynamic cell membrane remodelling. Nature 438:590-596.
6 - Chapman K.D., Ohlrogge J.B. (2012) Compartmentation of Triacylglycerol Accumulation in Plants. Journal of Biological Chemistry 287:2288-2294.
__________________________________________________________________________________________________________________
An Unusual Pathway for Saturated Fatty Acid Biosynthesis
There is at least one additional pathway for saturated fatty acid biosynthesis in plants. The main pathway discussed in lecture 19 is generally known as the fatty acid synthase (FAS) pathway. This is responsible for most medium chain fatty acid synthesis and all known long and very long chain fatty acid synthesis. Another pathway for elongation of short and medium length, branched and straight chain alkyl groups has been described by Kroumova and Wagner and designated the alpha-ketoacid elongation (a-KAE) pathway.
The a-KAE pathway appears to be mainly involved in formation of the alkyl components of trichome exudate (gum) sugar esters in Nicotianas and petunia. In contrast to the FAS cycle, which elongates by 2 C per cycle, the a-KAE pathway elongates by one C per cycle. The biochemistry of the a-KAE pathway corresponds to the formation of a-ketoglutarate from OAA in the TCA cycle and is derived from branched chain amino acid (bcAA; Val, Leu, Ileu) metabolism (shaded areas)(Fig. 1; Kroumova et al., 1994):
![[]](diag2.jpg)
A comparison of the predicted labeling patterns of products of the 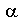 -KAE and FAS fed
[2-14C]acetate is shown in Fig. 2 of Kroumova et al.
(1994). Percentages of total label expected in the activated
carboxyl carbon are given under each molecule. Green, underlined
carbons are predicted to be labeled if synthesized via -KAE and carbons
with a blue asterisk via FAS:
-KAE and FAS fed
[2-14C]acetate is shown in Fig. 2 of Kroumova et al.
(1994). Percentages of total label expected in the activated
carboxyl carbon are given under each molecule. Green, underlined
carbons are predicted to be labeled if synthesized via -KAE and carbons
with a blue asterisk via FAS:
![[]](diag3.gif)
In Nicotiana 4-methyl-1-hexanol of headspace is synthesized via FAS and
the 4-methyl-hexanoyl-CoA acyl group of sucrose esters via -KAE (Kandra and Wagner.
1998).
Plant Membrane Lipid Biosynthesis
Membrane structure -- newer view Fig. 1 Engelman (2005).
Membrane curvature -- Fig. 1 and Fig. 3 McMahon and Gallop (2005).
The acyl groups of membrane and storage lipids (as well as cutin, suberin and waxes) are derived from FAS. As mentioned in Lecture 19, plant cell membranes usually contain significant amounts of unsaturated fatty acids. The main mono-unsaturated fatty acid formed in plants, oleic acid (18:1), is synthesized from stearoyl-ACP by a soluble stromal enzyme, stearoyl-ACP desaturase. All other unsaturated fatty acids in plants are synthesized by membrane bound desaturases. Plant cells have 2 pathways of unsaturated fatty acid and membrane glycerolipid biosynthesis, the so-called prokaryotic and eukaryotic pathways. The prokaryotic pathway occurs in the inner envelope of plastids and uses acyl-ACPs as substrates and the eukaryotic pathway in the cytosol (mainly ER) and uses acyl-CoAs as substrates. Glycerolipid synthesis begins with transfer of an acyl group, preferably 18:1, from ACP or CoA to glycerol-3-P (G3P) by glycerol-3-P acyltransferase (G3-P-AT or GPAT) producing lysophosphatidic acid (LPA) or monoacylglycerol-3-P. A fatty acid is transferred to the sn-2 position by LPA-AT or LPAT forming phosphatidic acid (PA). The fatty acid transferred by LPAT is predominately 16:0 in the prokaryotic pathway and an unsaturated fatty acid, usually 18:1 in the eukaryotic pathway. This difference can be used as a measure of the relative contributions of the prokaryotic and eukaryotic pathways in the formation of particular lipids. The PA is either converted to diacylglycerol by PA phosphatase or reacts with cytidine 5'-triphosphate (CTP) forming the nucleotide-activated form of DAG, CDP-DAG. This nucleotide activation provides the energy for polar headgroup attachment. CDP-DAG reacts with myo-inositol, serine and G3P to form PI, PS and phosphatidyl glycerol phosphate (PG). In the synthesis of the other membrane lipids, an activated headgroup reacts with DAG. CDP-choline, CDP-ethanolamine, UDP-galactose and UDP-sulfoquinovose react with DAG forming PC, PE, MGD and SL. DGD is synthesized from MGD. This is illustrated in Fig. 4 of Ohlrogge and Browse (1995) The Plant Cell 7, 957-970 and Fig. 10.29 of the class text:
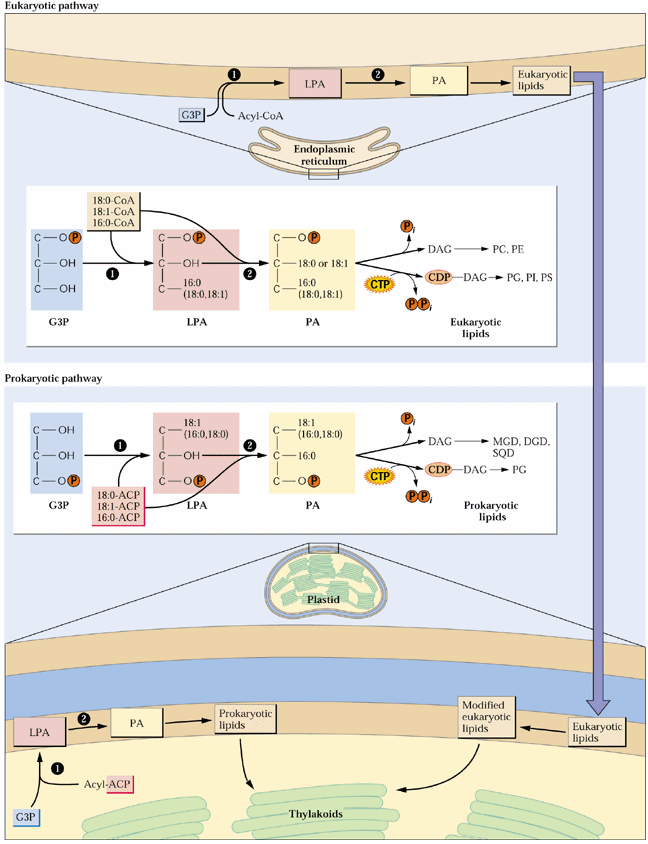
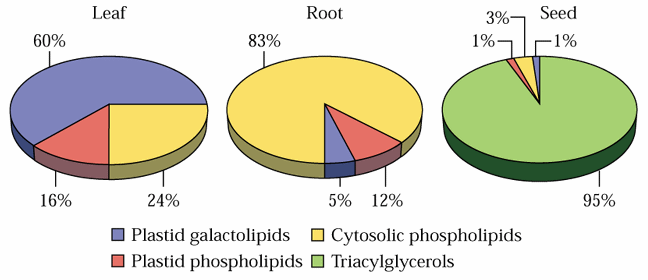
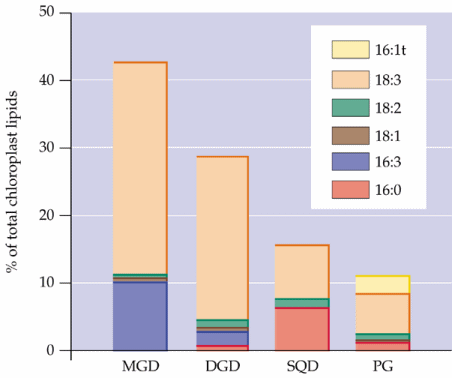
Chloroplasts of lower plants such as green algae are essentially autonomous in membrane lipid synthesis, but in the evolution of higher plants, an increasing proportion of plastid lipids are synthesized by the eukaryotic pathway. In many species of higher plants PG is the only lipid fully synthesized in plastids [see Fig. 5 of Ohlrogge and Browse (1995) and Fig. 10.21 of the class text below]. Different plant tissues vary widely in glycerolipid composition:
Fatty Acid Desaturation
Most of the membrane lipid fatty acids are further desaturated by membrane-bound desaturases. The contributions of different desaturases to this process have mostly been elucidated by analysis of genetic mutants by Browse and Somerville et al. Chloroplasts contain 4 membrane-bound desaturases. Two have a high degree of substrate specificity. One of these, a D3 desaturase (Fad4), inserts a trans double bond only into 16:0 at the sn-2 position of PG. The other (Fad5) places a cis double bond at the D7 position of 16:0 mainly at the sn-2 position of MGD and possibly DGD. The 3rd desaturase (Fad6) places a second cis double bond 3 carbons after the first double bond toward the w end of 16:1 or 18:1 at either position of MGD, DGD or SL. The 4th desaturase, for which there are 2 isozymes in Arabidopsis leaves (Fad7,8), inserts a cis double bond at the w3 position of 16:2 and 18:2. In most chloroplasts, w3 desaturases convert nearly all the di-unsaturated to tri-unsaturated fatty acids. In the ER oleic acid is desaturated to linoleic acid by an ER desaturase (Fad2) and linoleic acid can be further desaturated to linolenic acid (by Fad3). Plant membrane lipid desaturations are illustrated in Fig. 5 of Ohlrogge and Browse (1995) The Plant Cell 7, 957-970 and Fig. 10.21 of the class text:
![[]](Browsefig.jpg)
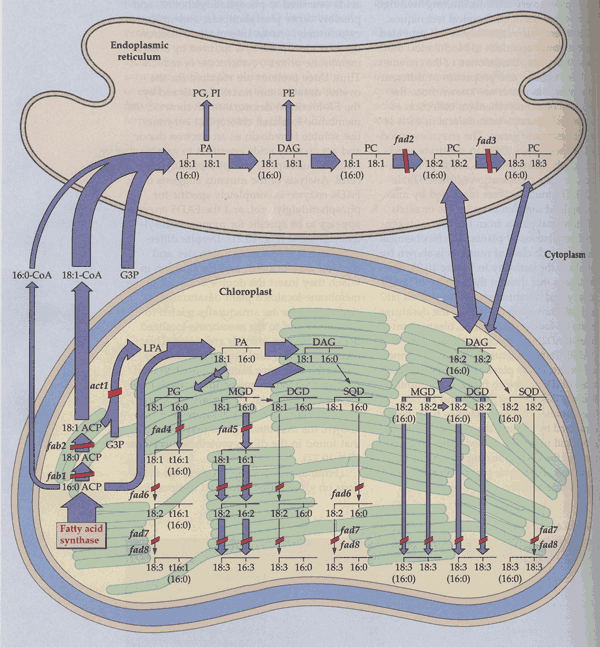
A number of plants can rather rapidly change their membrane lipid desaturation in response to temperature. This is a fundamental aspect of chilling and freezing as well as heat tolerance as described by Murakami et al. (2000). Plant lipids have tremendous differences in fatty acid compositions.
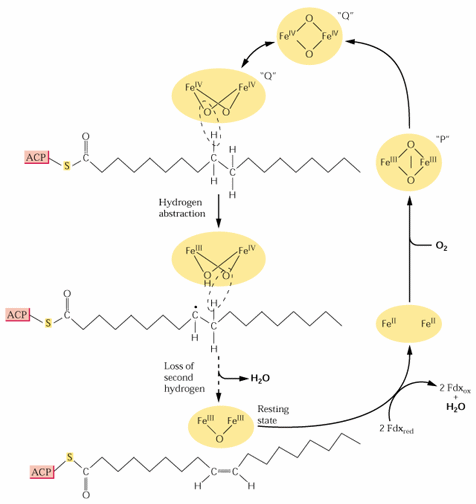
Fatty acid desaturases are nonheme di-iron oxidases that transfer 4 e-, 2 from the fatty acid substrate and 2 from a reductant, to O2 forming 2 H2Os. The ER desaturases require NADH, cytochrome b5 and cytochrome b5 reductase. The plastid desaturases apparently utilize reduced ferredoxin.
The ER D12 desaturase reaction is illustrated in the following figure:
![[]](desat.gif)
The structure of the soluble ACP desaturase has been determined and found to contain two proximal iron atoms per enzyme monomer.
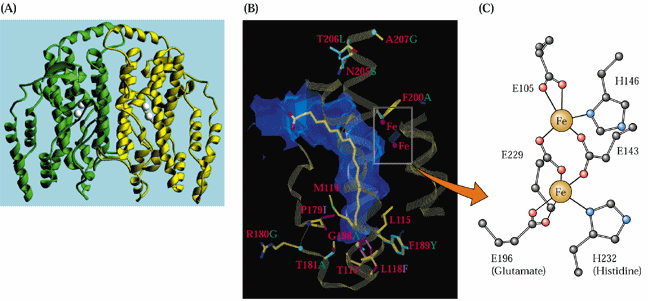
The desaturases are an interesting group of iron oxidases that must be reduced by an electron donor before oxidizing their fatty acid substrates.
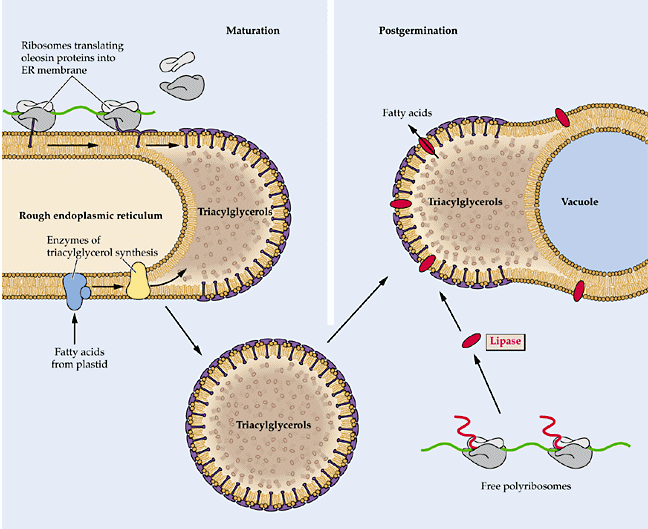
Triacylglycerol Biosynthesis
The main storage lipids in plants are the triacylglycerols (TAG). TAG is synthesized via the so-called Kennedy pathway. This pathway apparently operates in the ER and the TAG accumulates in structures know as oil bodies which are surrounded by a phospholipid membrane monolayer rather than the usual bilayer. The oil bodies contain one major type of proteins known as oleosins.
The Kennedy pathway involves 4 enzymatic steps involving 3 acyltransferases and a phosphatase. The 1st and 2nd acyltransferase reactions, which transfer fatty acids from fatty acid-CoAs to the sn-1 and sn-2 positions of glycerol, are common to membrane lipid synthesis (as is the phosphatase). The 3rd acyltransferase, diacylglycerol acyltransferase which esterifies a fatty acid at the sn-3 position, is unique to TAG biosynthesis:
![[]](kp.gif)
Renewable energy - World energy supplies, fuel...
Membrane lipids are composed mainly of the 5 fatty acids discussed previously. However, TAGs are known to accumulate >300 fatty acids/fatty acid derivatives. This is apparently due to a combination of unique fatty acid biosynthetic/modification enzymes in oil accumulating tissues and specificity of acyltransferases directing these unusual fatty acids from the acyl-CoA pool into TAG.
Two new routes for triglyceride biosynthesis have recently been discovered:
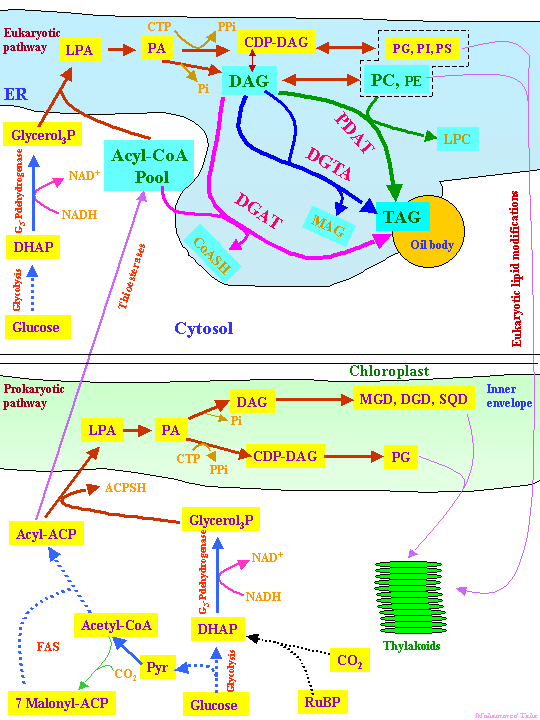
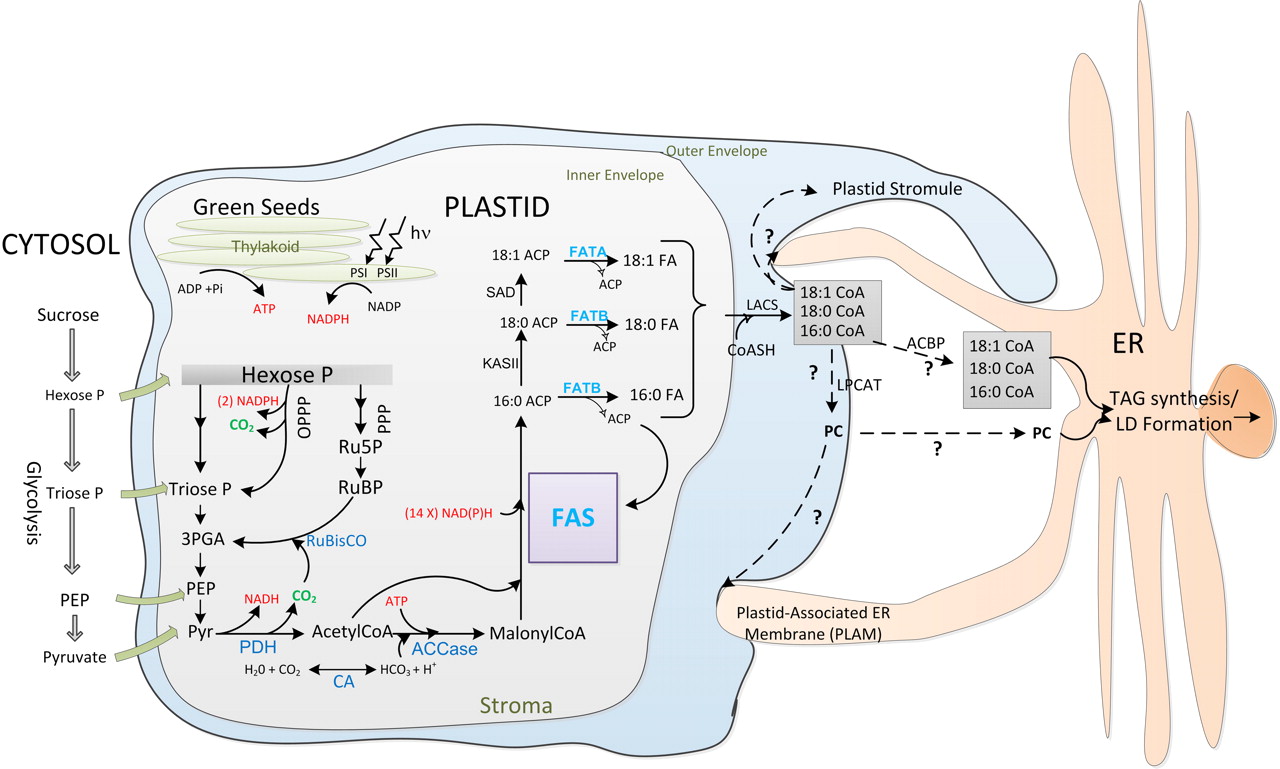
Figs. 1 & 2. of Chapman & Ohlrogge (2012) provides the latest understanding of TAG biosynthesis in plant tissues such as seeds:
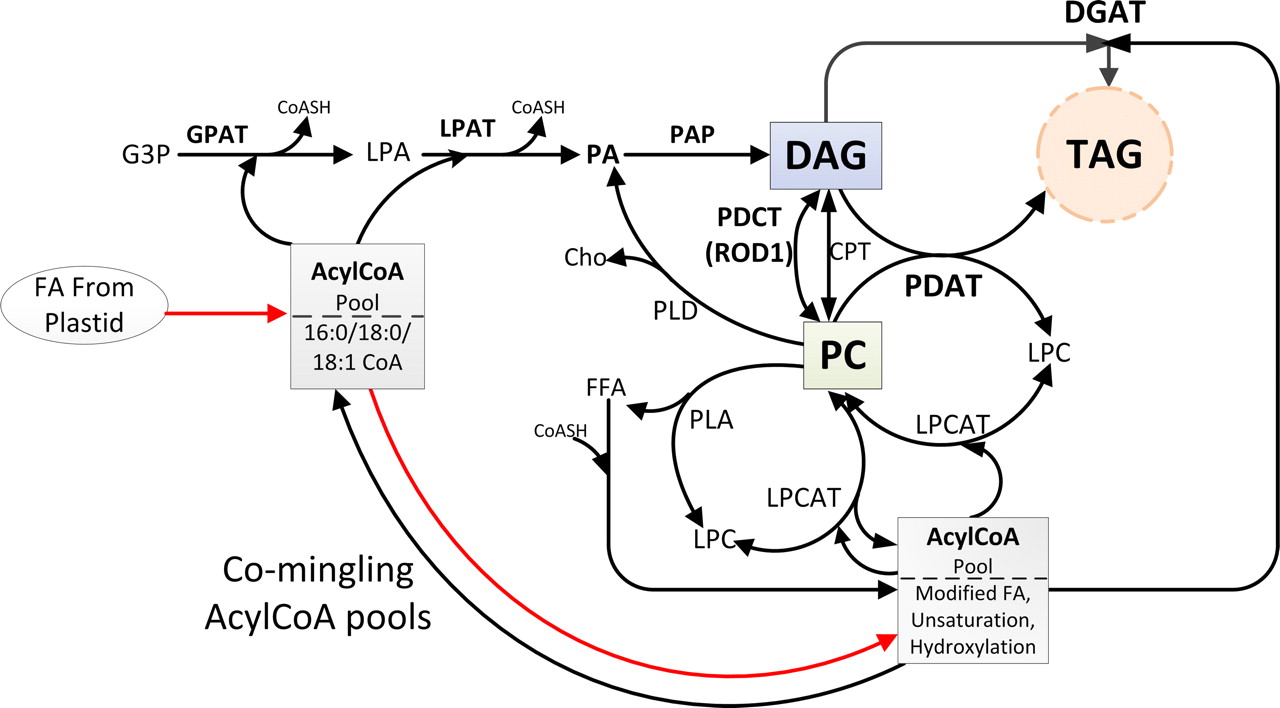
One way in which the diversity in fatty acids that can accumulate in TAG is achieved is by recruiting existing fatty acid biosynthetic enzymes to evolve new functions. This is illustrated by an altered D-12 desaturase which instead catalyzes hydroxylation instead of desaturation. Broun et al. (1998) were able to convert an oleate D-12 desaturase into a hydroxylase with as few as 4 amino acid substitutions and a hydroxylase into a D-12 desaturase with only 6 amino acid substitutions (Plant Journal 13:201-210)!
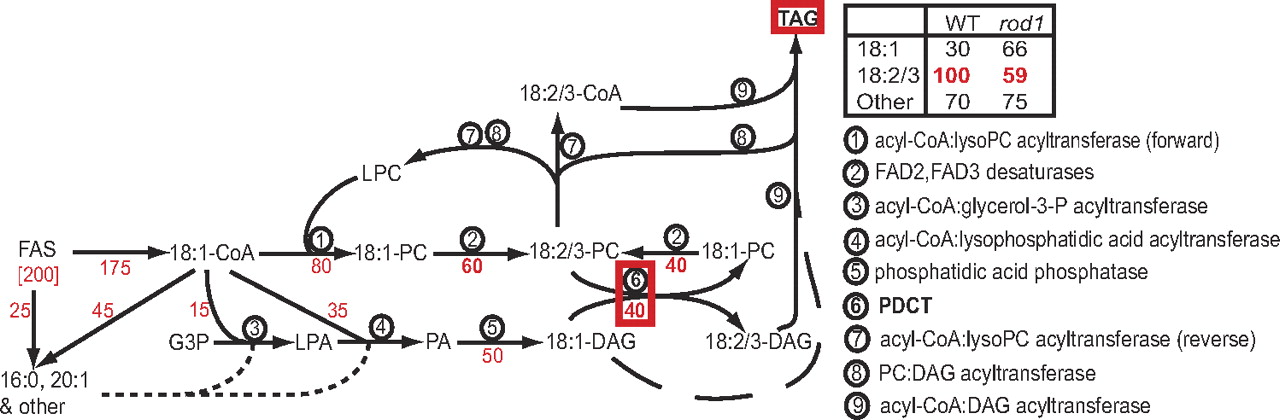
Lu et al. (2009) (PNAS 106:18837) describe a new enzyme, phosphatidylcholine:diacylglycerol cholinephosphotransferase (PDCT), that can retailor much of the DAG precurser for TAG biosynthesis. See Figure 5 of Lu et al. (2009).
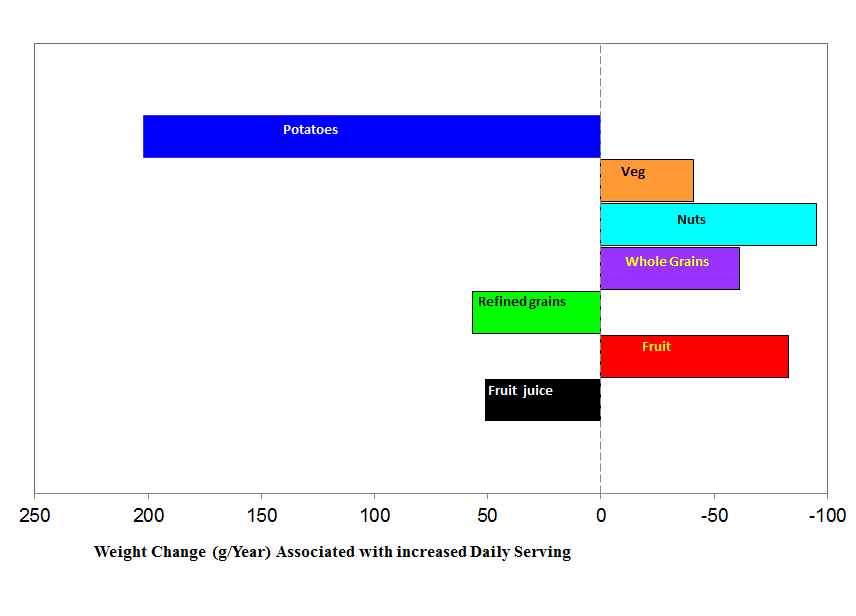
Mozaffarian, D., Hao, T., Rimm, E.B., Willett, W.C., and Hu, F.B. (2011). Changes in Diet and Lifestyle and Long-Term Weight Gain in Women and Men. New England Journal of Medicine 364, 2392-2404.
From light harvestion and CO2 fixation to accumulation of TAG in seeds poster
______________________________________________________________________________________________________________________
Reading Assignment for the Final Lipid Lecture Discussion:
a) REQUIRED:
1 - Chapter 10, sections 10.10.4 -10.10.5, 10.11 and Chapt. 17 section 17.8 of the Biochemistry & Molecular Biology of Plants class text.
b) OPTIONAL:
1 - Browse, J. 2009. Jasmonate Passes Muster: A Receptor and Targets for the Defense Hormone. Annual Review of Plant Biology 60:183-205.
2 - Koch, et al. 1999. Differential induction of plant volatile biosynthesis in the lima bean by early and late intermediates of the octadecanoid-signaling pathway. Plant Physiol. 121:153-162.
3 - Hamberg, M., A. Sanz, M. J. Rodriguez, A. P. Calvo and C. Castresana. 2003. Activation of the Fatty Acid {alpha}-Dioxygenase Pathway during Bacterial Infection of Tobacco Leaves: FORMATION OF OXYLIPINS PROTECTING AGAINST CELL DEATH. J. Biol. Chem. 278: 51796-51805.
4 - Lorenzo, O. R. Piqueras, J.J. Sanchez-Serrano and R. Solano. 2003. ETHYLENE RESPONSE FACTOR1 Integrates Signals from Ethylene and Jasmonate Pathways in Plant Defense. Plant Cell 15: 165-178.
5 - Pracharoenwattana, I., J.E. Cornah, and S.M. Smith. 2005. Arabidopsis Peroxisomal Citrate Synthase Is Required for Fatty Acid Respiration and Seed Germination. Plant Cell 17: 2037-2048.
__________________________________________________________________________________________________________________
| All materials © 2017 David Hildebrand, unless otherwise noted. | |||||||
| home | syllabus | lecture schedule & web notes | virtual office hours | messages & answers from the instructor | supplementary material | related links | What's new? |
{kind=link}
{kind=link}